본 명세서에 사용되는 바와 같이, 용어 "피브로넥틴 III형(FN3) 도메인"(FN3 도메인)은 피브로넥틴, 테나신, 세포내 세포골격 단백질, 사이토카인 수용체 및 원핵생물성 효소를 포함하는 단백질에서 자주 나타나는 도메인을 지칭한다(문헌[Bork and Doolittle, Proc Nat Acad Sci USA 89: 8990-8994, 1992]; 문헌[Meinke et al., J Bacteriol 175:1910-1918, 1993]; 문헌[Watanabe et al., J Biol Chem 265:15659-15665, 1990]).예시적인 FN3 도메인은 인간 테나신 C에 존재하는 15개의 상이한 FN3 도메인, 인간 피브로넥틴(FN)에 존재하는 15개의 상이한 FN3 도메인, 및 예를 들어 미국 특허 출원 공개 제2010/0216708호에 기재된 바와 같은 비천연 합성 FN3 도메인이다. 개별적인 FN3 도메인은 도메인 번호 및 단백질 명칭(예를 들어, 테나신의 제3 FN3 도메인(TN3), 또는 피브로넥틴의 제10 FN3 도메인(FN10))에 의해 지칭된다.
본 명세서에 사용되는 바와 같이, 용어 "치환하는" 또는 "치환된" 또는 "돌연변이화하는" 또는 "돌연변이화된"은 폴리펩티드 또는 폴리뉴클레오티드 서열 내에 그 서열의 변이체를 생성시키기 위해 하나 이상의 아미노산 또는 뉴클레오티드를 변경, 결실, 또는 삽입하는 것을 지칭한다.
본 명세서에 사용되는 바와 같이, 용어 "무작위화하는" 또는 "무작위화된" 또는 "다양화된(diversified)" 또는 "다양화하는"은 폴리뉴클레오티드 또는 폴리펩티드 서열 내에 적어도 하나의 치환, 삽입, 또는 결실을 실행하는 것을 지칭한다.
본 명세서에 사용되는 바와 같이, "변이체"는 하나 이상의 변형(modification), 예를 들어 치환, 삽입, 또는 결실에 의해 기준 폴리펩티드 또는 기준 폴리뉴클레오티드와 상이한 폴리펩티드 또는 폴리뉴클레오티드를 지칭한다.
본 명세서에 사용되는 바와 같이, 용어 "특이적으로 결합한다" 또는 "특이적 결합"은 본 발명의 FN3 도메인이 일정 해리 상수(KD)를 갖고서 사전결정된 항원에 결합하는 능력을 지칭하는데, 이때 해리 상수는 1×10-6 M 이하, 예를 들어 1×10-7 M 이하, 1×10-8 M 이하, 1×10-9 M 이하, 1×10-10 M 이하, 1×10-11 M 이하, 1×10-12 M 이하, 또는 1×10-13 M 이하이다. 전형적으로, 본 발명의 FN3 도메인은 예를 들어 프로테온(Proteon) 기기(바이오라드(BioRad))를 사용하여 표면 플라즈몬 공명(surface plasmon resonance)에 의해 측정할 때 비특이적 항원(예를 들어, BSA 또는 카세인)에 대한 그의 KD보다 적어도 10배 더 적은 KD를 갖고서 사전결정된 항원(즉, EGFR 또는 c-Met)에 결합한다. 따라서, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 EGFR 및 c-Met 둘 모두에 대해 적어도 1×10-6 M 이하의 결합 친화도(KD)를 갖고서 각각의 EGFR 및 c-Met에 특이적으로 결합한다. 그러나, 사전결정된 항원에 특이적으로 결합하는 본 발명의 단리된 FN3 도메인은 다른 관련 항원들과의, 예를 들어 다른 종으로부터의 동일한 사전결정된 항원(상동체)과의 교차-반응성을 가질 수 있다.
용어 "라이브러리"는 변이체들의 수집을 지칭한다. 라이브러리는 폴리펩티드 또는 폴리뉴클레오티드 변이체들로 구성될 수 있다.
본 명세서에 사용되는 바와 같이, 용어 "안정성"은 분자가 그의 정상적인 기능적 활성들 중 적어도 하나, 예를 들어 EGFR 또는 c-Met와 같은 사전결정된 항원에 결합하는 활성을 유지하도록, 생리학적 조건 하에서 접힌 상태를 유지하는 분자의 능력을 지칭한다.
본 명세서에 사용되는 바와 같이, "표피 성장 인자 수용체" 또는 "EGFR"은 서열 번호 73에 그리고 진뱅크(GenBank) 기탁 번호 NP_005219에 나타낸 서열뿐만 아니라 이의 천연 발생 변이체도 갖는 인간 EGFR(HER-1 또는 Erb-B1로도 알려짐)(문헌[Ullrich et al., Nature 309:418-425, 1984])을 지칭한다. 그러한 변이체는 잘 알려진 EGFRvIII 및 다른 대안적으로 스플라이싱된 변이체(예를 들어, 스위스프로트(SwissProt) 기탁 번호 P00533-1, P00533-2, P00533-3, P00533-4에 의해 확인된 바와 같음), 변이체 GLN-98, ARG-266, Lys-521, ILE-674, GLY-962, 및 PRO-988(문헌[Livingston et al., NIEHS-SNPs, environmental genome project, NIEHS ES15478])을 포함한다.
본 명세서에 사용되는 바와 같이, "EGFR 리간드"는 EGFR에 대한 모든(예를 들어, 생리학적) 리간드를 포함하는데, 이는 EGF, TGF-α, 헤파린 결합 EGF(HB-EGF), 암피레귤린(amphiregulin, AR), 및 에피레귤린(epiregulin, EPI)을 포함한다.
본 명세서에 사용되는 바와 같이, "표피 성장 인자"(EGF)는 서열 번호 74에 나타낸 아미노산 서열을 갖는 잘 알려진 53 아미노산 인간 EGF를 지칭한다.
본 명세서에 사용되는 바와 같이, "간세포 성장 인자 수용체" 또는 "c-Met"는 서열 번호 101에 또는 진뱅크 기탁 번호 NP_001120972에 나타낸 아미노산 서열 및 이의 천연 변이체를 갖는 인간 c-Met를 지칭한다.
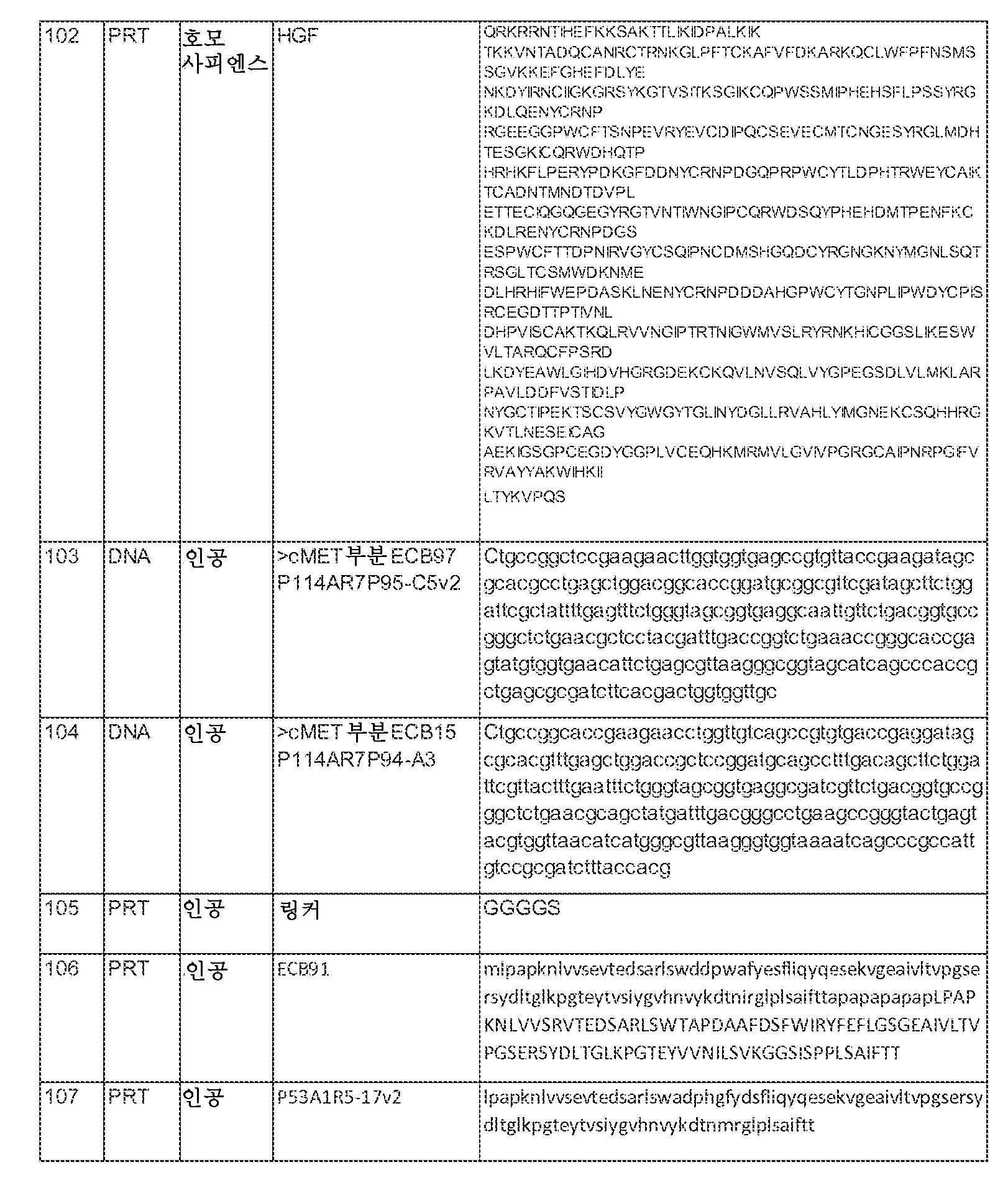
본 명세서에 사용되는 바와 같이, "간세포 성장 인자"(HGF)는 서열 번호 102에 나타낸 아미노산 서열을 갖는 잘 알려진 인간 HGF를 지칭하는데, 이는 절단되어서 이황화물 결합에 의해 연결된 알파 및 베타 쇄의 이량체를 형성한다.
본 명세서에 상호교환적으로 사용되는 바와 같이, "결합을 차단한다" 또는 "결합을 억제한다"는, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 본 발명의 FN3 도메인이 EGFR에 대한 EGFR 리간드, 예컨대 EGF의 결합 및/또는 c-Met에 대한 HGF의 결합을 차단 또는 억제하는 능력을 지칭하며, 부분 차단/억제 및 완전 차단/억제 둘 모두를 포함한다. 본 발명의 FN3 도메인 또는 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에 의한 EGFR 리간드, 예컨대 EGF의 EGFR에 대한 그리고/또는 HGF의 c-Met에 대한 차단/억제는, 차단 또는 억제를 갖지 않는 EGFR에 대한 EGFR 리간드 결합 및/또는 c-Met에 대한 HGF의 결합과 비교할 때, EGFR 신호전달 및/또는 c-Met 신호전달의 정상 수준을 부분적으로 또는 완전히 감소시킨다. 본 발명의 FN3 도메인 또는 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는, 억제율이 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%인 경우, EGFR 리간드, 예컨대 EGF의 EGFR에 대한 그리고/또는 HGF의 c-Met에 대한 "결합을 차단한다". 결합의 억제율은 잘 알려진 방법을 사용하여 측정될 수 있는데, 예를 들어, FACS를 사용하고 본 명세서에 기재된 방법을 사용하여 본 발명의 FN3 도메인 또는 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에 노출된 EGFR 발현 A431 세포 상에의 비오티닐화 EGF의 결합의 억제를 측정함으로써 측정되거나, 또는 잘 알려진 방법 및 본 명세서에 기재된 방법을 사용하여 c-Met 세포외 도메인 상에의 비오티닐화 HGF의 결합의 억제를 측정함으로써 측정될 수 있다.
용어 "EGFR 신호전달"은 EGFR에 결합하는 EGFR 리간드에 의해 유도되는 신호 전달(signal transduction)을 지칭하는데, 이러한 신호 전달은 EGFR 내의 적어도 하나의 티로신 잔기의 자가인산화로 이어진다. 예시적인 EGFR 리간드는 EGF이다.
본 명세서에 사용되는 바와 같이, "EGFR 신호전달을 무력화한다(neutralize)"는 것은 본 발명의 FN3 도메인이 EGFR 리간드, 예컨대 EGF에 의해 유도되는 EGFR 신호전달을 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 억제하는 능력을 지칭한다.
용어 "c-Met 신호전달"은 c-Met에 결합하는 HGF에 의해 유도되는 신호 전달을 지칭하는데, 이러한 신호 전달은 c-Met 내의 적어도 하나의 티로신 잔기의 자가인산화로 이어진다. 전형적으로, 위치 1230, 1234, 또는 1235의 적어도 하나의 티로신 잔기가 HGF 결합시에 자가인산화된다.
본 명세서에 사용되는 바와 같이, "c-Met 신호전달을 무력화한다"는 것은 본 발명의 FN3 도메인이 HGF에 의해 유도되는 c-Met 신호전달을 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 억제하는 능력을 지칭한다.
본 명세서에 상호교환적으로 사용되는 바와 같이, "과발현", "과발현된" 및 "과발현하는"은 동일한 조직 유형의 정상 세포와 비교하여 표면 상에 측정가능할 정도로 더 높은 수준의 EGFR 및/또는 c-Met를 갖는 암 또는 악성 세포를 지칭한다. 그러한 과발현은 유전자 증폭에 의해 또는 증가된 전사 또는 번역에 의해 야기될 수 있다. EGFR 및/또는 c-Met의 발현 및 과발현은 잘 알려진 검정을 사용하여, 예를 들어 살아 있거나 또는 용해된 세포에 대해 ELISA, 면역형광, 유세포측정 또는 방사면역검정을 사용하여 측정될 수 있다. 대안적으로 또는 추가적으로, EGFR 및/또는 c-Met-암호화 핵산 분자의 수준이, 예를 들어 형광 계내 혼성화(in situ hybridization), 서던 블롯팅(Southern blotting), 또는 PCR 기법을 사용하여 세포에서 측정될 수 있다. 세포의 표면 상에서의 EGFR 및/또는 c-Met의 수준이 정상 세포와 비교할 때 적어도 1.5배 초과인 경우, EGFR 및/또는 c-Met는 과발현된다.
본 명세서에 사용되는 바와 같이, "텐콘"은 서열 번호 1에 나타내고 미국 특허 출원공개US2010/0216708호에 기재된 서열을 갖는 합성 피브로넥틴 III형(FN3) 도메인을 지칭한다.
본 명세서에 사용되는 바와 같이, "암 세포" 또는 "종양 세포"는 생체내(in vivo)에서의, 생체외(ex vivo)에서의, 그리고 조직 배양물에서의 암성, 전-암성(pre-cancerous) 또는 형질전환된 세포를 지칭하며, 이러한 세포는 자발적 또는 유도된 표현형 변화를 갖는데, 이러한 변화는 반드시 새로운 유전 물질의 흡수를 수반하지는 않는다. 형질전환이 형질전환 바이러스에 의한 감염 및 새로운 게놈 핵산의 도입, 또는 외인성 핵산의 흡수로부터 일어날 수 있기는 하지만, 이는 또한 자발적으로 또는 발암물질에 대한 노출 후에 일어날 수 있으며, 그럼으로써 내인성 유전자를 돌연변이화할 수 있다. 형질전환/암은, 예를 들어, 시험관내에서, 생체내에서, 그리고 생체외에서 누드 마우스 등과 같은 적합한 동물 숙주에서 형태학적 변화, 세포의 불멸화, 비정상 성장 제어, 병소 형성, 증식, 악성종양, 종양 특이적 마커 수준, 침습성, 종양 성장 또는 억제에 의해 예시된다(문헌[Freshney, Culture of Animal Cells: A Manual of Basic Technique (3rd ed. 1994)]).
용어 "벡터"는 생물 시스템 내에서 복제될 수 있거나 그러한 시스템들 사이에서 이동할 수 있는 폴리뉴클레오티드를 의미한다. 전형적으로 벡터 폴리뉴클레오티드는 생물 시스템에서 이들 폴리뉴클레오티드의 복제 또는 유지를 용이하게 하는 기능을 하는 복제 기점, 폴리아데닐화 신호 또는 선택 마커와 같은 요소들을 포함한다. 그러한 생물 시스템의 예는 벡터를 복제할 수 있는 생물 구성요소들을 이용하는 세포 시스템, 바이러스 시스템, 동물 시스템, 식물 시스템 및 재구성된 생물 시스템을 포함할 수 있다. 벡터를 포함하는 폴리뉴클레오티드는 DNA 또는 RNA 분자 또는 이들의 혼성체일 수 있다.
용어 "발현 벡터"는 발현 벡터 내에 존재하는 폴리뉴클레오티드 서열에 의해 암호화되는 폴리펩티드의 번역을 유도하기 위하여 생물 시스템 또는 재구성된 생물 시스템에서 이용될 수 있는 벡터를 의미한다.
용어 "폴리뉴클레오티드"는 당-포스페이트 골격 또는 다른 등가의 공유결합적 화학적 특성에 의해 공유 결합된 뉴클레오티드의 쇄를 포함하는 분자를 의미한다. 이중 가닥 및 단일 가닥 DNA 및 RNA가 폴리뉴클레오티드의 전형적인 예이다.
용어 "폴리펩티드" 또는 "단백질"은 폴리펩티드를 형성하도록 펩티드 결합에 의해 연결된 적어도 2개의 아미노산 잔기를 포함하는 분자를 의미한다. 약 50개 미만의 아미노산의 작은 폴리펩티드는 "펩티드"로 지칭될 수 있다.
본 명세서에 사용되는 바와 같이, 용어 "이중특이성 EGFR/c-Met 분자" 또는 "이중특이성 EGFR/c-Met FN3 도메인 함유 분자"는 직접적으로 또는 링커를 통해 함께 공유결합적으로 연결된, EGFR 결합 FN3 도메인과 별개의 c-Met 결합 FN3 도메인을 포함하는 분자를 지칭한다. 예시적인 이중특이성 EGFR/c-Met 결합 분자는 EGFR과 특이적으로 결합하는 제1 FN3 도메인 및 c-Met와 특이적으로 결합하는 제2 FN3 도메인을 포함한다.
본 명세서에 사용되는 바와 같이, "결합가(valent)"는 분자 내의 항원에 특이적인 명시된 수의 결합 부위의 존재를 지칭한다. 그렇기 때문에, 용어 "1가", "2가", "4가", 및 "6가"는 분자 내의 항원에 특이적인 각각 1개, 2개, 4개, 및 6개의 결합 부위의 존재를 지칭한다.
본 명세서에 사용되는 바와 같이, "혼합물"은 함께 공유결합적으로 연결되지 않은 2개 이상의 FN3 도메인의 샘플 또는 조제물을 지칭한다. 혼합물은 2개 이상의 동일한 FN3 도메인 또는 별개의 FN3 도메인으로 이루어질 수 있다.
대상 조성물
본 발명은 시스테인 조작된 단일특이성 및 이중특이성 EGFR 및/또는 c-Met 결합 FN3 도메인 함유 분자 및 그의 제조 및 사용 방법을 제공한다.
단일특이성 EGFR 결합 분자
본 발명은 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하며, 이에 따라 치료적 및 진단적 응용에 널리 사용될 수 있는 피브로넥틴 III형(FN3) 도메인을 제공한다. 본 발명은 본 발명의 FN3 도메인을 암호화하는 폴리뉴클레오티드 또는 그의 상보적 핵산, 벡터, 숙주 세포, 및 이들의 제조 및 사용 방법을 제공한다.
본 발명의 FN3 도메인은 고 친화도로 EGFR에 결합하여 EGFR 신호전달을 억제하며, 소분자 EGFR 억제제와 비교할 때 특이성 및 감소된 탈표적 독성의 측면에서, 그리고 종래의 항체 치료제와 비교할 때 개선된 조직 침투의 측면에서 이득을 제공할 수 있다.
본 발명의 일 실시 형태는 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하는 단리된 피브로넥틴 III형(FN3) 도메인이다.
본 발명의 FN3 도메인은, A431 세포를 채용하고 본 발명의 FN3 도메인과 함께 또는 이것 없이 인큐베이션된 A431 세포 상에서 600 nM에서 스트렙타비딘-피코에리트린 접합체를 사용하여 결합된 비오티닐화 EGF로부터의 형광의 양을 검출하는 경쟁 검정에서, 약 1×10-7 M 미만, 약 1×10-8 M 미만, 약 1×10-9 M 미만, 약 1×10-10 M 미만, 약 1×10-11 M 미만, 또는 약 1×10-12 M 미만의 IC50 값으로 EGFR에 대한 EGF의 결합을 차단할 수 있다. 예시적인 FN3 도메인은 약 1×10-9 M 내지 약 1×10-7 M의 IC50 값으로 EGFR에 대한 EGF의 결합을 차단할 수 있으며, 예컨대 서열 번호 18-29, 107-110, 또는 122-137의 아미노산 서열을 갖는 EGFR 결합 FN3 도메인이다. 본 발명의 FN3 도메인은, 동일한 검정 조건을 사용하여 본 발명의 FN3 도메인의 부재 하에서의 EGFR에 대한 EGF의 결합과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 EGFR에 대한 EGF의 결합을 차단할 수 있다.
본 발명의 FN3 도메인은, 동일한 검정 조건을 사용하여 본 발명의 FN3 도메인의 부재 하에서의 신호전달의 수준과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 EGFR 신호전달을 억제할 수 있다.
EGFR에 대한 리간드, 예컨대 EGF의 결합은 수용체 이량체화, 자가인산화, 수용체의 내부 세포질 티로신 키나제 도메인의 활성화, 및 DNA 합성(유전자 활성화) 및 세포 주기 진행 또는 분열의 조절에 관여하는 다수의 신호 전달 및 전사활성화(transactivation) 경로들의 개시를 자극한다. EGFR 신호전달의 억제는 하나 이상의 EGFR 하류 신호전달 경로의 억제를 가져올 수 있으며, 이에 따라 EGFR을 무력화하는 것은 세포 증식 및 분화, 혈관생성, 세포 운동성 및 전이의 억제를 포함하는 다양한 효과를 가질 수 있다.
EGFR 신호전달은 잘 알려진 다양한 방법을 사용하여 측정될 수 있는데, 예를 들어 티로신 Y1068, Y1148, 및 Y1173 중 임의의 것에서의 수용체의 자가인산화(문헌[Downward et al., Nature 311:483-5, 1984]) 및/또는 천연 또는 합성 기질의 인산화를 측정하여 측정될 수 있다. 인산화는 포스포티로신 특이적 항체를 사용하여 ELISA 검정 또는 웨스턴 블롯과 같은 잘 알려진 방법을 사용하여 검출될 수 있다. 예시적인 검정은 문헌[Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997] 및 문헌[Batley et al., Life Sci 62:143-50, 1998]에서 찾을 수 있다.
일 실시 형태에서, 본 발명의 FN3 도메인은 50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 2.5×10-6 M 미만, 예를 들어 약 1×10-6 M 미만, 약 1×10-7 M 미만, 약 1×10-8 M 미만, 약 1×10-9 M 미만, 약 1×10-10 M 미만, 약 1×10-11 M 미만, 또는 약 1×10-12 M 미만의 IC50 값으로 EGFR 잔기 위치 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제한다.
일 실시 형태에서, 본 발명의 FN3 도메인은 50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 1.8 × 10-8 M 내지 약 2.5 × 10-6 M의 IC50 값으로 EGFR 잔기 위치 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제한다. 그러한 예시적인 FN3 도메인은 서열 번호 18-29, 107-110, 또는 122-137의 아미노산 서열을 갖는 것들이다.
일 실시 형태에서, 본 발명의 FN3 도메인은, 당업자에 의해 실시되는 바와 같이, 표면 플라즈몬 공명 또는 키넥사 방법(Kinexa method)에 의해 결정할 때, 약 1×10-8 M 미만, 예를 들어 약 1×10-9 M 미만, 약 1×10-10 M 미만, 약 1×10-11 M 미만, 약 1×10-12 M 미만, 또는 약 1×10-13 M 미만의 해리 상수(KD)를 갖고서 인간 EGFR에 결합한다. 일부 실시 형태에서, 본 발명의 FN3 도메인은 약 2×10-10 내지 약 1×10-8 M의 KD를 갖고서 인간 EGFR에 결합한다. EGFR에 대한 FN3 도메인의 친화도는 임의의 적합한 방법을 사용하여 실험적으로 결정될 수 있다.(예를 들어, 문헌[Berzofsky, et al., "Antibody-Antigen Interactions," In Fundamental Immunology, Paul, W. E., Ed., Raven Press: New York, NY (1984)]; 문헌[Kuby, Janis Immunology, W. H. Freeman and Company: New York, NY (1992)]; 및 본 명세서에 기재된 방법 참조). 특정 FN3 도메인-항원 상호작용의 측정된 친화도는 상이한 조건(예를 들어, 삼투성, pH) 하에서 측정된다면 변동될 수 있다. 따라서, 친화도 및 기타 항원-결합 파라미터(예를 들어, KD, Kon, Koff)의 측정은 바람직하게는 단백질 스캐폴드 및 항원의 표준화된 용액, 및 표준화된 완충액, 예컨대 본 명세서에 기재된 완충액을 사용하여 행해진다.
EGFR에 결합하는 본 발명의 예시적인 FN3 도메인은 서열 번호 18-29, 107-110, 또는 122-137의 FN3 도메인을 포함한다.
일 실시 형태에서, EGFR에 특이적으로 결합하는 FN3 도메인은 서열 번호 27의 아미노산 서열과 87% 이상 동일한 아미노산 서열을 포함한다.
일 실시 형태에서, EGFR에 특이적으로 결합하는 FN3 도메인은
서열 HNVYKDTNX9RGL(서열 번호 179) 또는 서열 LGSYVFEHDVML(서열 번호 180)(여기서, X9는 M 또는 I임)을 포함하는 FG 루프; 및
서열 X1X2X3X4X5X6X7X8(서열 번호 181)
(여기서,
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L임)을 포함하는 BC 루프를 포함한다.
EGFR에 특이적으로 결합하여 EGFR의 자가인산화를 억제하는 본 발명의 FN3 도메인은, 구조적 특징으로서, 서열 HNVYKDTNX9RGL(서열 번호 179) 또는 서열 LGSYVFEHDVML(서열 번호 180)(여기서, X9는 M 또는 I임)을 포함하는 FG 루프를 포함할 수 있다. 그러한 FN3 도메인은, 8개 또는 9개의 아미노산 길이를 갖고 서열 X1X2X3X4X5X6X7X8(서열 번호 181)에 의해 규정된 BC 루프를 추가로 포함하여, 50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 2.5×10-6 M 미만의 IC50 값으로, 그리고 약 1.8×10-8 M 내지 약 2.5×10-6 M의 IC50 값으로 EGFR 자가인산화를 억제할 수 있다.
EGFR에 특이적으로 결합하여 EGFR의 자가인산화를 억제하는 본 발명의 FN3 도메인은 서열
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX9RGLPLSAEFTT(서열 번호 182), 또는 서열 LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVLGSYVFEHDVMLPLSAEFTT(서열 번호 183)
(여기서,
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L이고;
X9는 M 또는 I임)를 추가로 포함한다.
EGFR 결합 FN3 도메인은 잘 알려진 방법 및 본 명세서에 기재된 방법을 사용하여 생성되고 EGFR 자가인산화를 억제하는 그의 능력에 대해 시험될 수 있다.
본 발명의 다른 실시 형태는 EGFR에 특이적으로 결합하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호 18-29, 107-110, 또는 122-137에 나타낸 서열을 포함한다.
일부 실시 형태에서, EGFR 결합 FN3 도메인은, 예를 들어 반감기 연장 분자의 발현 및/또는 접합을 용이하게 하기 위하여 특정 FN3 도메인의 N-말단에 연결된 개시자 메티오닌(Met) 또는 C-말단에 연결된 시스테인(Cys)을 포함한다.
본 발명의 다른 실시 형태는 EGFR에 특이적으로 결합하여 EGFR에 대한 EGF의 결합을 차단하는 단리된 피브로넥틴 III형(FN3) 도메인이며, 본 FN3 도메인은 서열 번호 1의 텐콘 서열에 기초하여 설계된 라이브러리로부터 단리된다.
단일특이성 c-Met 결합 분자
본 발명은 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하며, 이에 따라 치료적 및 진단적 응용에 널리 사용될 수 있는 피브로넥틴 III형(FN3) 도메인을 제공한다. 본 발명은 본 발명의 FN3 도메인을 암호화하는 폴리뉴클레오티드 또는 그의 상보적 핵산, 벡터, 숙주 세포, 및 이들의 제조 및 사용 방법을 제공한다.
본 발명의 FN3 도메인은 고 친화도로 c-Met에 결합하여 c-Met 신호전달을 억제하며, 소분자 c-Met 억제제와 비교할 때 특이성 및 감소된 탈표적 독성의 측면에서, 그리고 종래의 항체 치료제와 비교할 때 개선된 조직 침투의 측면에서 이득을 제공할 수 있다. 본 발명의 FN3 도메인은 1가이며, 이에 따라 다른 2가 분자들에서 일어날 수 있는 원치 않는 수용체 클러스터화 및 활성화를 방지한다.
본 발명의 일 실시 형태는 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하는, 단리된 피브로넥틴 III형(FN3) 도메인이다.
본 발명의 FN3 도메인은, 본 발명의 FN3 도메인의 존재 하에서의 c-Met-Fc 융합 단백질에 대한 비오티닐화 HGF의 결합의 억제를 검출하는 검정에서, 약 1×10-7 M 미만, 약 1×10-8 M 미만, 약 1×10-9 M 미만, 약 1×10-10 M 미만, 약 1×10-11 M 미만, 또는 약 1×10-12 M 미만의 IC50 값으로 c-Met에 대한 HGF의 결합을 차단할 수 있다. 예시적인 FN3 도메인은 약 2×10-10 M 내지 약 6×10-8의 IC50 값으로 c-Met에 대한 HGF의 결합을 차단할 수 있다. 본 발명의 FN3 도메인은, 동일한 검정 조건을 사용하여 본 발명의 FN3 도메인의 부재 하에서의 c-Met에 대한 HGF의 결합과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 c-Met에 대한 HGF의 결합을 차단할 수 있다.
본 발명의 FN3 도메인은, 동일한 검정 조건을 사용하여 본 발명의 FN3 도메인의 부재 하에서의 신호전달의 수준과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 c-Met 신호전달을 억제할 수 있다.
c-Met에 대한 HGF의 결합은 수용체 이량체화, 자가인산화, 수용체의 내부 세포질 티로신 키나제 도메인의 활성화, 및 DNA 합성(유전자 활성화) 및 세포 주기 진행 또는 분열의 조절에 관여하는 다수의 신호 전달 및 전사활성화 경로들의 개시를 자극한다. c-Met 신호전달의 억제는 하나 이상의 c-Met 하류 신호전달 경로의 억제를 가져올 수 있으며, 이에 따라 c-Met를 무력화하는 것은 세포 증식 및 분화, 혈관생성, 세포 운동성 및 전이의 억제를 포함하는 다양한 효과를 가질 수 있다.
c-Met 신호전달은 잘 알려진 다양한 방법을 사용하여 측정될 수 있는데, 예를 들어 적어도 하나의 티로신 잔기 Y1230, Y1234 또는 I1235 상에서의 수용체의 자가인산화, 및/또는 천연 또는 합성 기질의 인산화를 측정하여 측정될 수 있다. 인산화는, 예를 들어, ELISA 검정에서 또는 웨스턴 블롯 상에서 포스포티로신에 특이적인 항체를 사용하여 검출될 수 있다. 티로신 키나제 활성에 대한 몇몇 검정(문헌[Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997]; 문헌[Batley et al., Life Sci 62:143-50, 1998]).
일 실시 형태에서, 본 발명의 FN3 도메인은, 100 ng/mL의 재조합 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 1×10-6 M 미만, 약 1×10-7 M 미만, 약 1×10-8 M 미만, 약 1×10-9 M 미만, 약 1×10-10 M 미만, 약 1×10-11 M 미만, 또는 약 1×10-12 M 미만의 IC50 값으로 c-Met 잔기 위치 1349에서의 HGF-유도 c-Met 인산화를 억제한다.
일 실시 형태에서, 본 발명의 FN3 도메인은, 100 ng/mL의 재조합 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 4×10-9 M 내지 약 1×10-6 M의 IC50 값으로 c-Met 티로신 Y1349에서의 HGF-유도 c-Met 인산화를 억제한다.
일 실시 형태에서, 본 발명의 FN3 도메인은, 당업자에 의해 실시되는 바와 같이, 표면 플라즈몬 공명 또는 키넥사 방법에 의해 결정할 때, 약 1×10-7 M, 1×10-8 M, 1×10-9 M, 1×10-10 M, 1×10-11 M, 1×10-12 M, 1×10-13 M, 1×10-14 M, 또는 1×10-15 M 이하의 해리 상수(KD)를 갖고서 인간 c-Met에 결합한다.일부 실시 형태에서, 본 발명의 FN3 도메인은 약 3×10-10 내지 약 5×10-8 M의 KD를 갖고서 인간 c-Met에 결합한다. c-Met에 대한 FN3 도메인의 친화도는 임의의 적합한 방법을 사용하여 실험적으로 결정될 수 있다.(예를 들어, 문헌[Berzofsky, et al., "Antibody-Antigen Interactions," In Fundamental Immunology, Paul, W. E., Ed., Raven Press: New York, NY (1984)]; 문헌[Kuby, Janis Immunology, W. H. Freeman and Company: New York, NY (1992)]; 및 본 명세서에 기재된 방법 참조). 특정 FN3 도메인-항원 상호작용의 측정된 친화도는 상이한 조건(예를 들어, 삼투성, pH) 하에서 측정된다면 변동될 수 있다. 따라서, 친화도 및 기타 항원-결합 파라미터(예를 들어, KD, Kon, Koff)의 측정은 바람직하게는 단백질 스캐폴드 및 항원의 표준화된 용액, 및 표준화된 완충액, 예컨대 본 명세서에 기재된 완충액을 사용하여 행해진다.
c-Met에 결합하는 본 발명의 예시적인 FN3 도메인은 서열 번호 32-49 또는 111-114의 아미노산 서열을 갖는 FN3 도메인을 포함한다.
일 실시 형태에서, c-Met에 특이적으로 결합하는 FN3 도메인은 서열 번호 41의 아미노산 서열과 83% 이상 동일한 아미노산 서열을 포함한다.
일 실시 형태에서, c-Met에 특이적으로 결합하는 FN3 도메인은
서열 DSFX10IRYX11E X12X13X14X15GX16(서열 번호 184)
(여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D임)을 포함하는 C 가닥 및 CD 루프; 및
서열 TEYX17VX18IX19X20V KGGX21X22SX23(서열 번호 185)
(여기서,
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L임)을 포함하는 F 가닥 및 FG 루프를 포함한다.
c-Met에 특이적으로 결합하여 c-Met의 자가인산화를 억제하는 본 발명의 FN3 도메인은 서열:
LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX10IRYX11EX12X13X14X15GX16AIVLTVPGSERSYDLTGLKPGTEYX17VX18IX19X20VKGGX21X22SX23PLSAEFTT(서열 번호 186)
(여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D이고;
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L임)를 추가로 포함한다.
본 발명의 다른 실시 형태는 c-Met에 특이적으로 결합하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호 32-49 또는 111-114에 나타낸 서열을 포함한다.
본 발명의 다른 실시 형태는 c-Met에 특이적으로 결합하여 c-Met에 대한 HGF의 결합을 차단하는 단리된 피브로넥틴 III형(FN3) 도메인이며, 본 FN3 도메인은 서열 번호 1의 텐콘 서열에 기초하여 설계된 라이브러리로부터 단리된다.
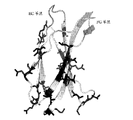
텐콘 서열에 기초한 라이브러리로부터의 EGFR 또는 c-Met FN3 도메인의 단리
텐콘(서열 번호 1)은 인간 테나신-C로부터의 15개의 FN3 도메인의 공통 서열로부터 설계된 비천연 발생 피브로넥틴 III형(FN3) 도메인이다(문헌[Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012]; 미국 특허 출원 공개제2010/0216708호). 텐콘의 결정 구조는 FN3 도메인들에 특징적인 바와 같은 7개의 베타-가닥을 연결시키는 6개의 표면-노출된 루프를 보여주며, 이때 베타-가닥은 A, B, C, D, E, F, 및 G로 지칭되고, 루프는 AB, BC, CD, DE, EF, 및 FG 루프로 지칭된다(문헌[Bork and Doolittle, Proc Natl Acad Sci USA 89:8990-8992, 1992]; 미국 특허 제6,673,901호). 이들 루프, 또는 각각의 루프 내의 선택된 잔기들은 EGFR에 결합하는 신규한 분자들을 선택하는 데 사용될 수 있는 피브로넥틴 III형(FN3) 도메인의 라이브러리를 작제하기 위하여 무작위화될 수 있다. 표 1은 텐콘(서열 번호 1) 내의 각각의 루프 및 베타-가닥의 위치 및 서열을 나타낸다.
따라서, 텐콘 서열에 기초하여 설계된 라이브러리는 무작위화된 FG 루프, 또는 무작위화된 BC 및 FG 루프를 가질 수 있으며, 예컨대 하기에 기재된 바와 같은 라이브러리 TCL1 또는 TCL2이다. 텐콘 BC 루프는 7개의 아미노산 길이이며, 이에 따라 1개, 2개, 3개, 4개, 5개, 6개 또는 7개의 아미노산이, BC 루프에서 다양화되고 텐콘 서열에 기초하여 설계된 라이브러리에서 무작위화될 수 있다. 텐콘 FG 루프는 7개의 아미노산 길이이며, 이에 따라 1개, 2개, 3개, 4개, 5개, 6개 또는 7개의 아미노산이, FG 루프에서 다양화되고 텐콘 서열에 기초하여 설계된 라이브러리에서 무작위화될 수 있다. 텐콘 라이브러리 내의 루프들에서의 추가의 다양성(diversity)이 루프들에서의 잔기들의 삽입 및/또는 결실에 의해 달성될 수 있다. 예를 들어, FG 및/또는 BC 루프는 1 내지 22개의 아미노산만큼 연장되거나, 또는 1 내지 3개의 아미노산만큼 감소될 수 있다. 텐콘 내의 FG 루프는 7개의 아미노산 길이인 반면에, 항체 중쇄 내의 상응하는 루프는 4 내지 28개 잔기의 범위이다. 최대 다양성을 제공하기 위하여, FG 루프는 서열에 있어서 뿐만 아니라 4 내지 28개 잔기의 항체 CDR3 길이 범위에 상응하도록 길이에 있어서도 다양화될 수 있다. 예를 들어, FG 루프는 1개, 2개, 3개, 4개 또는 5개의 추가 아미노산에 의해 루프를 연장시킴으로써 길이에 있어서 추가로 다양화될 수 있다.
텐콘 서열에 기초하여 설계된 라이브러리는 또한, FN3 도메인의 일측에 형성되고 2개 이상의 베타 가닥과 적어도 하나의 루프를 포함하는 무작위화된 대체 표면들을 가질 수 있다. 하나의 그러한 대체 표면은 C 및 F 베타-가닥과 CD 및 FG 루프 내의 아미노산들에 의해 형성된다(C-CD-F-FG 표면). 텐콘 대체 C-CD-F-FG 표면에 기초하여 설계된 라이브러리가 도 4에 나타나 있으며, 그러한 라이브러리들의 생성에 관한 상세한 설명이 미국 특허 출원 제13/852,930호에 기재되어 있다.
텐콘 서열에 기초하여 설계된 라이브러리는 또한 텐콘 변이체들, 예컨대 잔기 위치 11, 14, 17, 37, 46, 73, 또는 86(잔기 넘버링은 서열 번호 1에 상응함)에서 치환을 갖는, 개선된 열 안정성을 나타내는 텐콘 변이체들에 기초하여 설계된 라이브러리들을 포함한다. 예시적인 텐콘 변이체가 미국 특허 출원 공개 제2011/0274623호에 기재되어 있으며, 서열 번호 1의 텐콘과 비교할 때 치환 E11R, L17A, N46V, E86I를 갖는 텐콘27(서열 번호 99)을 포함한다.
[표 1]
텐콘 및 다른 FN3 서열 기반 라이브러리들은, 무작위 또는 규정된 세트의 아미노산들을 사용하여, 선택된 잔기 위치들에서 무작위화될 수 있다. 예를 들어, 20개의 천연 발생 아미노산 모두를 암호화하는 NNK 코돈을 사용하여 무작위 치환을 갖는 라이브러리 내의 변이체를 생성할 수 있다. 다른 다양화 스킴(scheme)에서는, DVK 코돈을 사용하여 아미노산 Ala, Trp, Tyr, Lys, Thr, Asn, Lys, Ser, Arg, Asp, Glu, Gly, 및 Cys를 암호화할 수 있다. 대안적으로, NNS 코돈을 사용하여 20개의 아미노산 잔기 모두를 발생시킴과 동시에 정지 코돈의 빈도를 감소시킬 수 있다. 예를 들어, 슬로노믹스(Slonomics)® 기술(http:_//www_sloning_com)을 사용하여, 다양화하고자 하는 위치에 편향된 아미노산 분포를 가진 FN3 도메인의 라이브러리를 합성할 수 있다. 이 기술은, 수천 개의 유전자 합성 과정에 충분한 보편적 구성 요소(building block)로서 작용하는 사전제조된 이중 가닥 트리플렛(pre-made double stranded triplet)들의 라이브러리를 사용한다. 트리플렛 라이브러리는 임의의 원하는 DNA 분자를 구성하는 데 필요한 모든 가능한 서열 조합을 나타낸다. 코돈 표기는 잘 알려진 IUB 코드에 따른다.
EGFR 또는 c-Met에 특이적으로 결합하는 본 발명의 FN3 도메인은, cis 디스플레이를 사용하여 스캐폴드 단백질들을 암호화하는 DNA 단편들을 RepA를 암호화하는 DNA 단편에 연결하여, 시험관내 번역 후에 형성되는 단백질-DNA 복합체들의 풀(pool) - 여기서, 각각의 단백질은 그것을 암호화하는 DNA와 안정하게 회합됨 - 을 생성함으로써 텐콘 라이브러리와 같은 FN3 라이브러리를 생성하고(미국 특허 제7,842,476호; 문헌[Odegrip et al., Proc Natl Acad Sci U S A 101, 2806-2810, 2004]), 당업계에 알려지고 실시예에 기재된 임의의 방법에 의해 EGFR 및/또는 c-Met에 대한 특이적 결합에 대해 라이브러리를 검정함으로써 단리될 수 있다. 사용될 수 있는 예시적인 잘 알려진 방법은 ELISA, 샌드위치 면역검정, 및 경쟁적 및 비경쟁적 검정이다(예를 들어, 문헌[Ausubel et al., eds, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley & Sons, Inc., New York] 참조). EGFR 또는 c-Met에 특이적으로 결합하는 확인된 FN3 도메인은, 본 명세서에 기재된 방법을 사용하여, EGFR에 대한 EGFR 리간드, 예컨대 EGF의 결합, 또는 c-Met에 대한 HGF의 결합을 차단하는 그의 능력에 대해, 그리고 EGFR 및/또는 c-Met 신호전달을 억제하는 그의 능력에 대해 추가로 특성화된다.
EGFR 또는 c-Met에 특이적으로 결합하는 본 발명의 FN3 도메인은, 템플릿으로서 임의의 FN3 도메인을 사용하여 라이브러리를 생성하고, 내부에 제공된 방법을 사용하여 EGFR 또는 c-Met에 특이적으로 결합하는 분자들을 위해 라이브러리를 스크리닝함으로써 생성될 수 있다. 사용될 수 있는 예시적인 FN3 도메인은 테나신 C의 제3 FN3 도메인(TN3)(서열 번호 75), 피브콘(Fibcon)(서열 번호 76), 및 피브로넥틴의 제10 FN3 도메인(FN10)(서열 번호 77)이다. 표준 클로닝 및 발현 기법을 사용하여 라이브러리를 벡터 내로 클로닝하거나 또는 라이브러리의 이중 가닥 cDNA 카세트를 합성하거나, 라이브러리를 시험관내에서 번역하거나, 또는 발현시킨다. 예를 들어, 리보솜 디스플레이(문헌[Hanes and Pluckthun, Proc Natl Acad Sci USA, 94, 4937-4942, 1997]), mRNA 디스플레이(문헌[Roberts and Szostak, Proc Natl Acad Sci USA, 94, 12297-12302, 1997]), 또는 다른 무세포(cell-free) 시스템(미국 특허 제5,643,768)이 사용될 수 있다. FN3 도메인 변이체의 라이브러리는, 예를 들어 임의의 적합한 박테리오파지의 표면 상에 디스플레이되는 융합 단백질로서 발현될 수 있다. 박테리오파지의 표면 상에 융합 폴리펩티드를 디스플레이하는 방법은 잘 알려져 있다(미국 특허 출원 공개 제2011/0118144호; 국제 특허 공개 WO2009/085462호; 미국 특허 제6,969,108호; 미국 특허 제6,172,197호; 미국 특허 제5,223,409호; 미국 특허 제6,582,915호; 미국 특허 제6,472,147호).
EGFR 또는 c-Met에 특이적으로 결합하는 본 발명의 FN3 도메인은 그의 특성을 개선하도록, 예컨대 열 안정성 및 열 접힘과 풀림의 가역성을 개선하도록 변형될 수 있다. 단백질 및 효소의 겉보기 열 안정성을 증가시키기 위하여 몇몇 방법이 적용되어 왔으며, 이는 고도로 유사한 열 안정성 서열들에 대한 비교를 기반으로 하는 합리적인 설계, 이황화물 가교를 안정화하는 설계, 알파-나선 성향을 증가시키는 돌연변이, 염 가교의 조작(engineering), 단백질의 표면 전하의 변경, 지향 진화(directed evolution), 및 공통 서열들의 조성을 포함한다(문헌[Lehmann and Wyss, Curr Opin Biotechnol, 12, 371-375, 2001]). 높은 열 안정성은 발현 단백질의 수율을 증가시키고, 용해도 또는 활성을 개선하고, 면역원성을 감소시키고, 제조 중에 콜드 체인(cold chain)의 필요성을 최소화할 수 있다. 텐콘(서열 번호 1)의 열 안정성을 개선하기 위해 치환될 수 있는 잔기는 잔기 위치 11, 14, 17, 37, 46, 73, 또는 86이며, 미국 특허 출원 공개 제2011/0274623호에 기재되어 있다. 이들 잔기에 상응하는 치환이 본 발명의 FN3 도메인 또는 이중특이성 FN3 도메인 함유 분자에 도입될 수 있다.
본 발명의 다른 실시 형태는 EGFR에 특이적으로 결합하여 EGFR에 대한 EGF의 결합을 차단하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호 18-29, 107-110, 122-137에 나타낸 서열을 포함하며, 텐콘(서열 번호 1) 내의 위치 11, 14, 17, 37, 46, 73, 및 86에 상응하는 하나 이상의 잔기 위치에서의 치환을 추가로 포함한다.
본 발명의 다른 실시 형태는 c-Met에 특이적으로 결합하여 c-Met에 대한 HGF의 결합을 차단하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호 32-49 또는 111-114에 나타낸 서열을 포함하고, 텐콘(서열 번호 1) 내의 위치 11, 14, 17, 37, 46, 73, 및 86에 상응하는 하나 이상의 잔기 위치에서의 치환을 추가로 포함한다.
예시적인 치환은 치환 E11N, E14P, L17A, E37P, N46V, G73Y 및 E86I(서열 번호 1에 따른 넘버링)이다.
일부 실시 형태에서, 본 발명의 FN3 도메인은 텐콘(서열 번호 1) 내의 치환 L17A, N46V, 및 E86I에 상응하는 치환을 포함한다.
EGFR에 특이적으로 결합하는 FN3 도메인(도 1)은 텐콘(서열 번호 1)과 비교할 때 연장된 FG 루프를 갖는다. 따라서, 텐콘(서열 번호 1) 내의 잔기 11, 14, 17, 37, 46, 73, 및 86에 상응하는 잔기는, 서열 번호 24의 FN3 도메인 - 여기서는, BC 루프 내에의 하나의 아미노산의 삽입으로 인해 상응하는 잔기가 잔기 11, 14, 17, 38, 74, 및 92임 - 을 제외하고는, 도 1a 및 도 1b에 나타낸 EGFR FN3 도메인 내의 잔기 11, 14, 17, 37, 46, 73 및 91이다.
본 발명의 다른 실시 형태는 EGFR에 특이적으로 결합하여 EGFR에 대한 EGF의 결합을 차단하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호18-29, 107-110, 또는 122-137에 나타낸 아미노산 서열을 포함하며, 텐콘(서열 번호 1) 내의 치환 L17A, N46V, 및 E86I에 상응하는 치환을 선택적으로 갖는다.
본 발명의 다른 실시 형태는 c-Met에 특이적으로 결합하여 c-Met에 대한 HGF의 결합을 차단하는 단리된 FN3 도메인이며, 본 FN3 도메인은 서열 번호 32-49 또는 111-114에 나타낸 아미노산 서열을 포함하며, 텐콘(서열 번호 1) 내의 치환 L17A, N46V, 및 E86I에 상응하는 치환을 선택적으로 갖는다.
단백질 안정성 및 단백질 불안정성의 측정은 단백질 완전성(integrity)의 동일하거나 상이한 양상이라고 볼 수 있다. 단백질은 열에 의해, 자외 또는 이온화 방사선, 액체 용액 중에 있는 경우에는 주위 삼투성 및 pH의 변화, 작은 기공-크기 여과에 의해 부과되는 기계적 전단력, 자외 방사선, 이온화 방사선, 예컨대 감마 조사에 의한 이온화 방사선, 화학적 탈수 또는 열 탈수, 또는 단백질 구조 붕괴를 야기할 수 있는 임의의 다른 작용 또는 힘에 의해 야기되는 변성에 민감하거나 또는 "불안정"하다. 분자의 안정성은 표준 방법을 사용하여 결정될 수 있다. 예를 들어, 표준 방법을 사용하여 열 용융("TM") 온도 - 분자의 절반이 풀리게 되는 섭씨 도(℃) 단위의 온도 - 를 측정함으로써 분자의 안정성을 결정할 수 있다. 전형적으로, TM이 높을수록 분자가 더 안정하다. 열에 부가하여, 화학적 환경 또한 단백질이 특정 3 차원 구조를 유지하는 능력을 변화시킨다.
일 실시 형태에서, EGFR 또는 c-Met에 결합하는 본 발명의 FN3 도메인은, TM의 증가에 의해 측정할 때, 조작 전의 동일한 도메인과 비교하여 적어도 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 또는 95% 또는 그 이상으로 증가된 안정성을 나타낸다.
화학적 변성을 다양한 방법에 의해 마찬가지로 측정할 수 있다. 화학적 변성제는 구아니디늄 하이드로클로라이드, 구아니디늄 티오시아네이트, 우레아, 아세톤, 유기 용매(DMF, 벤젠, 아세토니트릴), 염(황산암모늄, 브롬화리튬, 염화리튬, 브롬화나트륨, 염화칼슘, 염화나트륨); 환원제(예를 들어, 다이티오트레이톨, 베타-메르캅토에탄올, 다이니트로티오벤젠, 및 수소화물, 예컨대 붕수소화나트륨), 비이온성 및 이온성 세제, 산(예를 들어, 염산(HCl), 아세트산(CH3COOH), 할로겐화 아세트산), 소수성 분자(예를 들어, 인지질), 및 표적화 변성제를 포함한다. 변성 정도의 정량화는 기능적 특성, 예컨대 표적 분자에 결합하는 능력의 손실에 의존할 수 있거나, 또는 물리화학적 특성, 예컨대 응집에 대한 경향, 이전에는 용매에 접근 불가능했던 잔기의 노출, 또는 이황화물 결합의 붕괴 또는 형성에 의해 행할 수 있다.
일 실시 형태에서, EGFR 또는 c-Met에 결합하는 본 발명의 FN3 도메인은, 화학적 변성제로서 구아니디늄 하이드로클로라이드를 사용함으로써 측정할 때, 조작 전의 동일한 스캐폴드와 비교하여 적어도 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 또는 95% 또는 그 이상으로 증가된 안정성을 나타낸다. 증가된 안정성은, 잘 알려진 방법을 사용하여, 증가하는 농도의 구아니딘 하이드로클로라이드로 처리시에 감소되는 트립토판 형광의 함수로서 측정될 수 있다.
예를 들어 결합가(valency)를 증가시킴으로써 표적 분자 결합의 결합력(avidity)을 증가시키기 위한 수단으로서, 또는 2개 이상의 상이한 표적 분자에 동시에 결합하는 이중특이성 또는 다중특이성 스캐폴드를 생성하기 위하여, 본 발명의 FN3 도메인을 단량체, 이량체, 또는 다량체로서 생성할 수 있다. 예를 들어, 아미노산 링커, 예를 들어 폴리-글리신, 글리신 및 세린, 또는 알라닌 및 프롤린을 함유하는 링커의 포함에 의해 단일특이성, 이중특이성, 또는 다중특이성 단백질 스캐폴드를 연결함으로써 이량체 및 다량체를 생성할 수 있다. 예시적인 링커는 (GS)2(서열 번호 78), (GGGGS)5(서열 번호 79), (AP)2(서열 번호 80), (AP)5(서열 번호 81), (AP)10(서열 번호 82), (AP)20(서열 번호 83), A(EAAAK)5AAA(서열 번호 84) 링커를 포함한다. 이량체들 및 다량체들은 N으로부터 C로의 방향으로 서로 연결될 수 있다. 폴리펩티드를 신규의 연결된 융합 폴리펩티드 내로 연결하기 위하여 천연 발생 펩티드 링커뿐만 아니라 인공 펩티드 링커를 사용하는 것에 대해서도 문헌에 잘 알려져 있다(문헌[Hallewell et al., J Biol Chem 264, 5260-5268, 1989]; 문헌[Alfthan et al., Protein Eng. 8, 725-731, 1995]; 문헌[Robinson & Sauer, Biochemistry 35, 109-116, 1996]; 미국 특허 제5,856,456호).
이중특이성 EGFR/c-Met 결합 분자
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 소분자 EGFR 억제제와 비교할 때 특이성 및 감소된 탈표적 독성의 측면에서, 그리고 종래의 항체 치료제와 비교할 때 개선된 조직 침투의 측면에서 이득을 제공할 수 있다. 본 발명은 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자가, EGFR-결합 FN3 도메인과 c-Met-결합 FN3 도메인의 혼합물과 비교할 때, 유의하게 개선된 상승적 억제 효과를 제공한다는 의외의 발견에 적어도 부분적으로 기초한다. 이러한 분자는 종양 침투 및 체류를 최대화하기 위하여 EGFR 및 c-Met 둘 모두에 대한 특이적 친화도에 맞추어질 수 있다.
본 발명의 일 실시 형태는, 제1 피브로넥틴 III형(FN3) 도메인 및 제2 FN3 도메인을 포함하며, 제1 FN3 도메인은 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하고, 제2 FN3 도메인은 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하는, 단리된 이중특이성 FN3 도메인 함유 분자이다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 본 발명의 임의의 EGFR-결합 FN3 도메인과 임의의 c-Met-결합 FN3 도메인을 직접적으로 또는 링커를 통해 공유결합적으로 연결함으로써 생성될 수 있다. 따라서, 이중특이성 분자의 제1 FN3 도메인은 EGFR-결합 FN3 도메인에 대해 전술된 바와 같은 특성들을 가질 수 있고, 이중특이성 분자의 제2 FN3 도메인은 c-Met-결합 FN3 도메인에 대해 전술된 바와 같은 특성들을 가질 수 있다.
일 실시 형태에서, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제1 FN3 도메인은, 50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 2.5×10-6 M 미만의 IC50 값으로 EGFR 잔기 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제하고, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제2 FN3 도메인은, 100 ng/mL의 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 1.5×10-6 M 미만의 IC50 값으로 c-Met 잔기 티로신 1349에서의 HGF-유도 c-Met 인산화를 억제한다.
다른 실시 형태에서, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제1 FN3 도메인은, 50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 1.8 × 10-8 M 내지 약 2.5×10-6 M의 IC50 값으로 EGFR 잔기 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제하고, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제2 FN3 도메인은, 100 ng/mL의 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 4×10-9 M 내지 약 1.5×10-6 M의 IC50 값으로 c-Met 잔기 티로신 1349에서의 HGF-유도 c-Met 인산화를 억제한다.
다른 실시 형태에서, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제1 FN3 도메인은 약 1×10-8 M 미만의 해리 상수(KD)를 갖고서 인간 EGFR에 결합하고, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 제2 FN3 도메인은 약 5×10-8 M 미만의 KD를 갖고서 인간 c-Met에 결합한다.
EGFR 및 c-Met 둘 모두에 결합하는 이중특이성 분자에서, 제1 FN3 도메인은 약 2×10-10 내지 약 1×10-8 M의 KD를 갖고서 인간 EGFR에 결합하고, 제2 FN3 도메인은 약 3×10-10 내지 약 5×10-8 M의 KD를 갖고서 인간 c-Met에 결합한다.
EGFR 및 c-Met에 대한 이중특이성 EGFR/c-Met 분자의 친화도는 단일특이성 분자에 대해 전술된 바와 같이 결정될 수 있다.
본 발명의 이중특이성 EGFR/c-Met 분자 내의 제1 FN3 도메인은, A431 세포를 채용하고 제1 FN3 도메인과 함께 또는 이것 없이 인큐베이션된 A431 세포 상에서 600 nM에서 스트렙타비딘-피코에리트린 접합체를 사용하여 결합된 비오티닐화 EGF로부터의 형광의 양을 검출하는 검정에서, 약 1×10-9 M 내지 약 1.5×10-7 M의 IC50 값으로 EGFR에 대한 EGF의 결합을 차단할 수 있다. 본 발명의 이중특이성 EGFR/c-Met 분자 내의 제1 FN3 도메인은, 동일한 검정 조건을 사용하여 제1 FN3 도메인의 부재 하에서의 EGFR에 대한 EGF의 결합과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 EGFR에 대한 EGF의 결합을 차단할 수 있다.
본 발명의 이중특이성 EGFR/c-Met 분자 내의 제2 FN3 도메인은, 제2 FN3 도메인의 존재 하에서의 c-Met-Fc 융합 단백질에 대한 비오티닐화 HGF의 결합의 억제를 검출하는 검정에서, 약 2×10-10 M 내지 약 6×10-8 M의 IC50 값으로 c-Met에 대한 HGF의 결합을 차단할 수 있다. 이중특이성 EGFR/c-Met 분자 내의 제2 FN3 도메인은, 동일한 검정 조건을 사용하여 제2 FN3 도메인의 부재 하에서의 c-Met에 대한 HGF의 결합과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 c-Met에 대한 HGF의 결합을 차단할 수 있다.
본 발명의 이중특이성 EGFR/c-Met 분자는, 동일한 검정 조건을 사용하여 본 발명의 이중특이성 EGFR/c-Met 분자의 부재 하에서의 신호전달의 수준과 비교할 때, 적어도 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%로 EGFR 및/또는 c-Met 신호전달을 억제할 수 있다.
EGFR 및 c-Met 신호전달은 단일특이성 분자에 대해 전술된 바와 같은 다양한 잘 알려진 방법을 사용하여 측정될 수 있다.
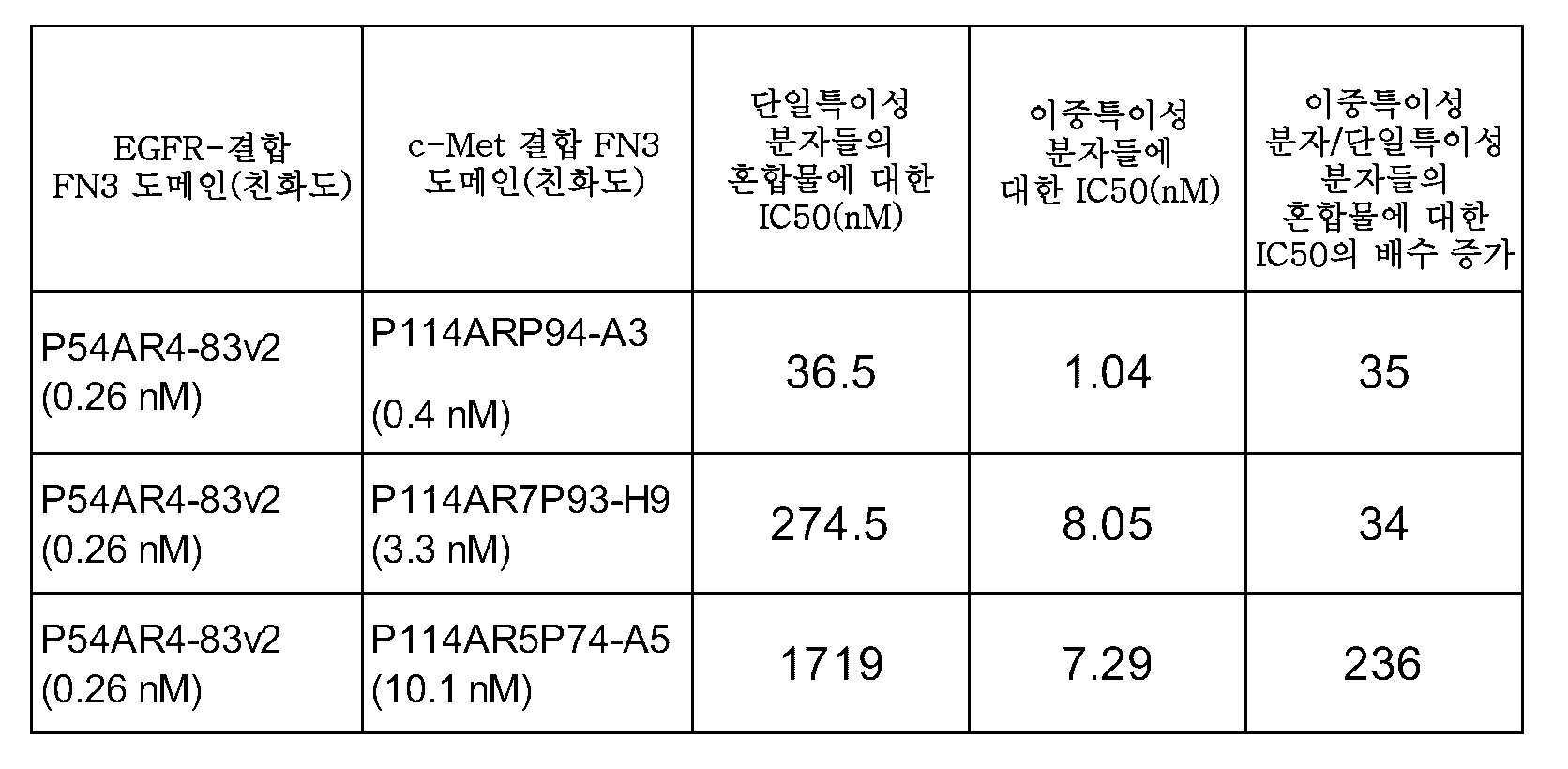
EGFR에 특이적으로 결합하는 제1 FN3 도메인 및 c-Met에 특이적으로 결합하는 제2 FN3 도메인을 포함하는 본 발명의 이중특이성 EGFR/c-Met 분자는, 제1 FN3 도메인과 제2 FN3 도메인의 혼합물에 의해 관찰된 상승적 억제와 비교할 때, EGFR 및 c/Met 신호전달 및 종양 세포 증식의 유의하게 증가된 상승적 억제를 제공한다. 상승적 억제는, 예를 들어 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에 의한 그리고 2개의 단일특이성 분자들 - 하나는 EGFR에 결합하고 다른 하나는 c-Met에 결합함 - 의 혼합물에 의한 ERK 인산화의 억제를 측정함으로써 평가될 수 있다. 본 발명의 이중특이성 EGFR/c-Met 분자는, 2개의 단일특이성 FN3 도메인들의 혼합물에 대한 IC50 값과 비교할 때, 적어도 약 100배 더 작은, 예를 들어 적어도 500, 1000, 5000 또는 10,000배 더 작은 IC50 값으로 ERK 인산화를 억제할 수 있는데, 이는 2개의 단일특이성 FN3 도메인들의 혼합물과 비교할 때 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에 대한 적어도 100배 증가된 효력을 나타낸다. 예시적인 이중특이성 EGFR-c-Met FN3 도메인 함유 분자는 약 5×10-9 M 이하의 IC50 값으로 ERK 인산화를 억제할 수 있다. ERK 인산화는 표준 방법 및 본 명세서에 기재된 방법을 사용하여 측정될 수 있다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는, 제1 FN3 도메인과 제2 FN3 도메인의 혼합물에 의한 H292 세포 성장의 억제의 IC50 값과 비교할 때 적어도 30배 더 적은 IC50 값으로 H292 세포 증식을 억제할 수 있으며, 여기서 세포 증식은 7.5 ng/mL의 HGF로 보충된 10% FBS를 함유하는 배지를 사용하여 유도된다.본 발명의 이중특이성 분자는, 제1 FN3 도메인과 제2 FN3 도메인의 혼합물에 의한 종양 세포 증식의 억제의 IC50 값과 비교할 때, 약 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300, 400, 500, 600, 700, 800, 또는 약 1000배 미만인 IC50 값으로 종양 세포 증식을 억제할 수 있다. 종양 세포 증식의 억제는 표준 방법 및 본 명세서에 기재된 방법을 사용하여 측정될 수 있다.
본 발명의 다른 실시 형태는, 제1 피브로넥틴 III형(FN3) 도메인 및 제2 FN3 도메인을 포함하며, 제1 FN3 도메인은 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하고, 제2 FN3 도메인은 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하는, 이중특이성 FN3 도메인 함유 분자이며, 여기서
제1 FN3 도메인은 서열 HNVYKDTNX9RGL(서열 번호 179) 또는 서열 LGSYVFEHDVML(서열 번호 180)(여기서, X9는 M 또는 I임)을 포함하는 FG 루프; 및
서열 X1X2X3X4X5X6X7X8(서열 번호 181)
(여기서,
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L임)을 포함하는 BC 루프를 포함하고;
제2 FN3 도메인은 서열 DSFX10IRYX11E X12X13X14X15GX16(서열 번호 184)
(여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D임)을 포함하는 C 가닥 및 CD 루프; 및
서열 TEYX17VX18IX19X20V KGGX21X22SX23(서열 번호 185)
(여기서,
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L임)을 포함하는 F 가닥 및 FG 루프를 포함한다.
다른 실시 형태에서, 이중특이성 분자는 서열:
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX9RGL PLSAEFTT(서열 번호 182), 또는 서열
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGV LGSYVFEHDVMLPLSAEFTT(서열 번호 183)를 포함하는, EGFR에 결합하는 제1 FN3 도메인을 포함하며,
여기서 서열 번호 X 및 X에서;
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L이고;
X9는 M 또는 I이다.
다른 실시 형태에서, 이중특이성 분자는 서열
LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX10IRYX11EX12X13X14X15GX16AIVLTVPGSERSYDLTGLKPGTEYX17VX18IX19X20VKGGX21X22SX23PLSAEFTT(서열 번호 186)를 포함하는, c-Met에 결합하는 제2 FN3 도메인을 포함하며,
여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D이고;
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L이다.
예시적인 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 서열 번호 50-72, 106, 118-121, 또는 138-165에 나타낸 아미노산 서열을 포함한다.
본 발명의 이중특이성 EGFR/c-Met 분자는 그의 기능적 특성, 예컨대 EGFR 자가인산화의 억제와 관련된 소정의 구조적 특성, 예컨대 서열HNVYKDTNX9RGL(서열 번호 179) 또는 서열 LGSYVFEHDVML(서열 번호 180)(여기서, X9는 M 또는 I임)을 포함하는, EGFR에 결합하는 제1 FN3 도메인의 FG 루프를 포함한다.
일 실시 형태에서, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는
50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 8×10-7 M 미만의 IC50 값으로 EGFR 잔기 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제하거나;
100 ng/mL의 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 8.4×10-7 M 미만의 IC50 값으로 c-Met 잔기 티로신 1349에서의 HGF-유도 c-Met 인산화를 억제하거나;
약 9.5×10-6 M 미만의 IC50 값으로 HGF-유도 NCI-H292 세포 증식을 억제하며, 여기서 세포 증식은 7.5 ng의 HGF를 함유하는 10% FBS를 사용하여 유도되거나;
약 2.0×10-8 M 미만의 KD를 갖고서 EGFR에 결합하거나;
약 2.0×10-8 M 미만의 KD를 갖고서 c-Met에 결합한다.
다른 실시 형태에서, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는
50 ng/mL의 인간 EGF를 사용하여 A431 세포에서 측정할 때, 약 4.2×10-9 M 내지 8×10-7 M의 IC50으로 EGFR 잔기 티로신 1173에서의 EGF-유도 EGFR 인산화를 억제하거나;
100 ng/mL의 인간 HGF를 사용하여 NCI-H441 세포에서 측정할 때, 약 2.4×10-8 M 내지 약 8.4×10-7 M의 IC50 값으로 c-Met 잔기 티로신 1349에서의 HGF-유도 c-Met 인산화를 억제하거나;
약 2.3×10-8 M 내지 약 9.5×10-6M의 IC50 값으로 HGF-유도 NCI-H292 세포 증식을 억제하며, 여기서 세포 증식은 7.5 ng의 HGF를 함유하는 10% FBS를 사용하여 유도되거나;
약 2×10-10 M 내지 약 2.0×10-8 M의 KD를 갖고서 EGFR에 결합하거나;
약 1×10-9 M 내지 약 2.0×10-8 M의 KD를 갖고서 c-Met에 결합한다.
일 실시 형태에서, 이중특이성 EGFR/c-Met 분자는 서열
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX9RGLPLSAEFTT(서열 번호 182)
(여기서,
X1은 D이고;
X2는 D이고;
X3은 P이고;
X4는 부재하고;
X5는 H 또는 W이고;
X6은 A이고;
X7은 F이고;
X8은 Y이고;
X9는 M 또는 I임)를 포함하는 EGFR-결합 FN3 도메인; 및
서열
PAPKNLVVSRVTEDSARLSWTAPDAAFDSFX10IRYX11EX12X13X14X15GX16AIVLTVPGSERSYDLTGLKPG TEYX17VX18IX19X20VKGGX21X22SX23PLSAEFTT(서열 번호 186)
(여기서,
X10은 W이고;
X11은 F이고;
X12는 F이고;
X13은 V 또는 L이고;
X14는 G 또는 S이고;
X15는 S 또는 K이고;
X16은 E 또는 D이고;
X17은 V이고;
X18은 N이고;
X19는 L 또는 M이고;
X20은 G 또는 S이고;
X21은 S 또는 K이고;
X22는 I이고;
X23은 P임)를 포함하는 c-Met-결합 FN3 도메인을 포함한다.
예시적인 이중특이성 EGFR/c-Met 분자는 서열 번호 57, 61, 62, 63, 64, 65, 66, 67 및 68에 나타낸 서열을 갖는 것들이다.
본 발명의 이중특이성 분자는, 전술된 바와 같은 텐콘(서열 번호 1) 내의 위치 11, 14, 17, 37, 46, 73 및 86에 상응하는 제1 FN3 도메인 및/또는 제2 FN3 도메인 내의 하나 이상의 잔기 위치에서의 치환, 및 위치 29에서의 치환을 추가로 포함할 수 있다. 예시적인 치환은 치환 E11N, E14P, L17A, E37P, N46V, G73Y, E86I 및 D29E(서열 번호 1에 따른 넘버링)이다. 당업자는 다른 아미노산들이 치환에 사용될 수 있음을 이해할 것이며, 이는 예컨대 하기에 기재된 바와 같은 측쇄에 관련된 아미노산 패밀리에 속하는 아미노산이다.생성된 변이체는 본 명세서에 있는 방법을 사용하여 그의 안정성 및 EGFR 및/또는 c-Met에 대한 그의 결합에 대해 시험될 수 있다.
일 실시 형태에서, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 EGFR에 특이적으로 결합하는 제1 FN3 도메인 및 c-Met에 특이적으로 결합하는 제2 FN3 도메인을 포함하며, 여기서 제1 FN3 도메인은 서열:
LPAPKNLVVSX24VTX25DSX26RLSWDDPX27AFYX28SFLIQYQX29SEKVGEAIX30LTVPGSERSYDLTGLKPGTEYTVSIYX31VHNVYKDTNX32RGLPLSAX33FTT(서열 번호 187)
(여기서,
X24는 E, N 또는 R이고;
X25는 E 또는 P이고;
X26은 L 또는 A이고;
X27은 H 또는 W이고;
X28은 E 또는 D이고;
X29는 E 또는 P이고;
X30은 N 또는 V이고;
X31은 G 또는 Y이고;
X32는 M 또는 I이고;
X33은 E 또는 I임)를 포함하고;
제2 FN3 도메인은 서열:
LPAPKNLVVSX34VTX35DSX36RLSWTAPDAAFDSFWIRYFX37FX38X39X40GX41AIX42LTVPGSERSYDLTGLKPGTEYVVNIX43X44VKGGX45ISPPLSAX46FTT(서열 번호 188)
(여기서,
X34는 E, N 또는 R이고;
X35는 E 또는 P이고;
X36은 L 또는 A이고;
X37은 E 또는 P이고;
X38은 V 또는 L이고;
X39는 G 또는 S이고;
X40은 S 또는 K이고;
X41은 E 또는 D이고;
X42는 N 또는 V이고;
X43은 L 또는 M이고;
X44는 G 또는 S이고;
X45는 S 또는 K이고;
X46은 E 또는 I임)를 포함한다.
다른 실시 형태에서, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 서열 번호 27의 아미노산 서열과 87% 이상 동일한 아미노산 서열을 포함하는 제1 FN3 도메인, 및 서열 번호 41의 아미노산 서열과 83% 이상 동일한 아미노산 서열을 포함하는 제2 FN3 도메인을 포함한다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 종양 축적률을 최대화하기 위해 EGFR 및 c-Met에 대한 특이적 친화도에 맞추어질 수 있다.
본 발명의 다른 실시 형태는 제1 피브로넥틴 III형(FN3) 도메인 및 제2 FN3 도메인을 포함하며, 제1 FN3 도메인은 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하고, 제2 FN3 도메인은 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하며, 여기서 제1 FN3 도메인 및 제2 FN3 도메인은 서열 번호 1의 텐콘 서열에 기초하여 설계된 라이브러리로부터 단리되는, 단리된 이중특이성 FN3 도메인 함유 분자이다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 잘 알려진 방법을 사용하여 본 발명의 EGFR-결합 FN3 도메인과 c-Met 결합 FN3 도메인을 공유결합적으로 커플링함으로써 생성될 수 있다. 이들 FN3 도메인은 링커, 예를 들어 폴리-글리신, 글리신 및 세린, 또는 알라닌 및 프롤린을 함유하는 링커를 통해 연결될 수 있다. 예시적인 링커는 (GS)2(서열 번호 78), (GGGGS)5(서열 번호 79), (AP)2(서열 번호 80), (AP)5(서열 번호 81), (AP)10(서열 번호 82), (AP)20(서열 번호 83), A(EAAAK)5AAA(서열 번호 84) 링커를 포함한다. 폴리펩티드를 신규의 연결된 융합 폴리펩티드 내로 연결하기 위하여 천연 발생 펩티드 링커뿐만 아니라 인공 펩티드 링커를 사용하는 것에 대해서도 문헌에 잘 알려져 있다(문헌[Hallewell et al., J Biol Chem 264, 5260-5268, 1989]; 문헌[Alfthan et al., Protein Eng. 8, 725-731, 1995]; 문헌[Robinson & Sauer, Biochemistry 35, 109-116, 1996]; 미국 특허 제5,856,456호). 본 발명의 이중특이성 EGFR/c-Met 분자는 제1 FN3 도메인의 C-말단으로부터 제2 FN3 도메인의 N-말단으로, 또는 제2 FN3 도메인의 C-말단으로부터 제1 FN3 도메인의 N-말단으로 함께 연결될 수 있다. 임의의 EGFR-결합 FN3 도메인은 c-Met-결합 FN3 도메인에 공유결합적으로 연결될 수 있다. 예시적인 EGFR-결합 FN3 도메인은 서열 번호 18-29, 107-110, 및 122-137에 나타낸 아미노산 서열을 갖는 도메인이고, 예시적인 c-Met 결합 FN3 도메인은 서열 번호 32-49 및 111-114에 나타낸 아미노산 서열을 갖는 도메인이다. 이중특이성 분자에 커플링하고자 하는 EGFR-결합 FN3 도메인은 그의 N-말단에서 개시자 메티오닌(Met)을 추가로 포함할 수 있다.
이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 변이체가 본 발명의 범주 내에 있다. 예를 들어, 생성된 변이체가 부모 분자와 비교할 때 EGFR 및 c-Met에 대한 유사한 선택성 및 효력을 유지하는 한, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에서 치환이 이루어질 수 있다. 예시적인 변형은, 예를 들어, 부모 분자의 특성과 유사한 특성을 갖는 변이체를 생성하게 될 보존적 치환이다. 보존적 대체는 측쇄에서 관련된 아미노산 패밀리 내에서 일어나는 것들이다. 유전적으로 암호화된 아미노산은 하기 4개의 패밀리로 나눌 수 있다: (1) 산성(아스파르테이트, 글루타메이트); (2) 염기성(라이신, 아르기닌, 히스티딘); (3) 비극성(알라닌, 발린, 류신, 아이소류신, 프롤린, 페닐알라닌, 메티오닌, 트립토판); 및 (4) 비하전 극성(글리신, 아스파라긴, 글루타민, 시스테인, 세린, 트레오닌, 티로신). 페닐알라닌, 트립토판, 및 티로신은 때때로 방향족 아미노산으로 공동으로 분류된다. 대안적으로, 아미노산 레퍼토리(repertoire)는 하기로 그룹화될 수 있다: (1) 산성(아스파르테이트, 글루타메이트); (2) 염기성(라이신, 아르기닌, 히스티딘), (3) 지방족(글리신, 알라닌, 발린, 류신, 아이소류신, 세린, 트레오닌) - 이때, 세린 및 트레오닌은 별도로 지방족-하이드록실로서 선택적으로 분류함 -; (4) 방향족(페닐알라닌, 티로신, 트립토판); (5) 아미드(아스파라긴, 글루타민); 및 (6) 황-함유(시스테인 및 메티오닌)(문헌[Stryer (ed.), Biochemistry, 2nd ed, WH Freeman and Co., 1981]). 이중특이성 EGFR/c-Met FN3 도메인 함유 분자에 대해 비보존적 치환이 이루어질 수 있는데, 비보존적 치환은 이중특이성 분자의 특성을 개선하기 위하여 상이한 아미노산 부류들 사이에서의 아미노산 잔기의 치환을 포함한다. 폴리펩티드 또는 그의 단편의 아미노산 서열의 변화가 기능적 상동체를 생성하는지는 본 명세서에 기재된 검정을 사용하여 변형 폴리펩티드 또는 단편이 비변형 폴리펩티드 또는 단편과 유사한 방식으로 반응을 생성하는 능력을 평가함으로써 용이하게 결정될 수 있다. 하나 초과의 대체가 일어난 펩티드, 폴리펩티드 또는 단백질은 동일한 방식으로 용이하게 시험될 수 있다.
예를 들어 결합가를 증가시킴으로써 표적 분자 결합의 결합력을 증가시키기 위한 수단으로서, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자를 이량체 또는 다량체로서 생성할 수 있다. 다량체는, 예를 들어 잘 알려진 방법을 사용하여 아미노산 링커의 포함에 의해, 하나 이상의 EGFR-결합 FN3 도메인과 하나 이상의 c-Met-결합 FN3 도메인을 연결하여, 적어도 3개의 개별 FN3 도메인을 포함하고 EGFR 또는 c-Met에 대해 적어도 이중특이성인 분자를 형성함으로써 생성될 수 있다.
본 발명의 다른 실시 형태는, 제1 피브로넥틴 III형(FN3) 도메인 및 제2 FN3 도메인을 포함하며, 제1 FN3 도메인은 표피 성장 인자 수용체(EGFR)에 특이적으로 결합하여 EGFR에 대한 표피 성장 인자(EGF)의 결합을 차단하고, 제2 FN3 도메인은 간세포 성장 인자 수용체(c-Met)에 특이적으로 결합하여 c-Met에 대한 간세포 성장 인자(HGF)의 결합을 차단하며, 서열 번호 50-72 또는 106에 나타낸 아미노산 서열을 포함하는, 이중특이성 FN3 도메인 함유 분자이다.
반감기 연장 모이어티(half-life extending moiety)
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자 또는 단일특이성 EGFR 또는 c-Met 결합 FN3 도메인은, 예를 들어 공유결합성 상호작용을 통해 다른 하위단위들을 포함할 수 있다. 본 발명의 일 태양에서, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 반감기 연장 모이어티를 추가로 포함한다. 예시적인 반감기 연장 모이어티는 알부민, 알부민-결합 단백질 및/또는 도메인, 트랜스페린 및 그의 단편 및 유사체, 및 Fc 영역이다. 예시적인 알부민-결합 도메인은 서열 번호 117에 나타나 있다.
항체 불변 영역의 전부 또는 일부를 본 발명의 분자에 부착하여 항체-유사 특성, 특히 Fc 영역과 관련된 특성, 예컨대 Fc 효과기 기능, 예컨대 C1q 결합, 보체 의존성 세포독성(complement dependent cytotoxicity, CDC), Fc 수용체 결합, 항체-의존성 세포-매개 세포독성(antibody-dependent cell-mediated cytotoxicity, ADCC), 식작용, 세포 표면 수용체(예를 들어, B 세포 수용체; BCR)의 하향 조절을 부여할 수 있으며, 이들 활성을 담당하는 Fc 내의 잔기를 변형시킴으로써 추가로 변형시킬 수 있다(검토를 위해서는 문헌[Strohl, Curr Opin Biotechnol.20, 685-691, 2009] 참조).
원하는 특성을 위해 추가 모이어티, 예컨대 폴리에틸렌 글리콜(PEG) 분자, 예컨대 PEG5000 또는 PEG20,000, 상이한 쇄 길이의 지방산 및 지방산 에스테르, 예를 들어 라우레이트, 미리스테이트, 스테아레이트, 아라키데이트, 베헤네이트, 올레에이트, 아라키도네이트, 옥탄이산, 테트라데칸이산, 옥타데칸이산, 도코산이산 등, 폴리라이신, 옥탄, 탄수화물(덱스트란, 셀룰로스, 올리고당 또는 다당)을 본 발명의 이중특이성 분자 내로 도입시킬 수 있다. 이들 모이어티는 단백질 스캐폴드 암호화 서열과의 직접 융합일 수 있으며, 표준 클로닝 및 발현 기법에 의해 생성될 수 있다. 대안적으로, 잘 알려진 화학적 커플링 방법을 사용하여 본 발명의 재조합적으로 생성된 분자에 이러한 모이어티를 부착할 수 있다.
예를 들어, 시스테인 잔기를 분자의 C-말단에 도입시키고 잘 알려진 방법을 사용하여 페길 기를 시스테인에 부착함으로써, 페길 모이어티를 본 발명의 이중특이성 또는 단일특이성 분자에 부가할 수 있다. C-말단 시스테인을 갖는 예시적인 이중특이성 분자는 서열 번호 170-178에 나타낸 아미노산 서열을 갖는 것들이다.
추가 모이어티를 도입시킨 본 발명의 단일특이성 및 이중특이성 분자는 몇몇 잘 알려진 검정에 의해 기능성에 대해 비교될 수 있다. 예를 들어, Fc 도메인 및/또는 Fc 도메인 변이체의 도입으로 인해 변경된 단일특이성 및/또는 이중특이성 분자의 특성은, Fc 수용체 결합 검정에서, 가용성 형태의 수용체, 예컨대 FcγRI, FcγRII, FcγRIII 또는 FcRn 수용체를 사용하여, 또는, 예를 들어 ADCC 또는 CDC를 측정하거나 또는 생체내 모델에서 본 발명의 분자의 약동학적 특성을 평가하는 잘 알려진 세포-기반 검정을 사용하여 검정될 수 있다.
폴리뉴클레오티드, 벡터, 숙주 세포
본 발명은 단리된 폴리뉴클레오티드로서 또는 발현 벡터의 일부분으로서 또는 선형 DNA 서열의 일부분으로서 본 발명의 EGFR-결합 또는 c-Met-결합 FN3 도메인 또는 이중특이성 EGFR/c-Met FN3 도메인 함유 분자를 암호화하는 핵산을 제공하며, 이때 선형 DNA 서열은 시험관내 전사/번역에 사용되는 선형 DNA 서열, 즉 그러한 조성 또는 그의 유도 돌연변이원의 원핵생물성, 진핵생물성 또는 사상(filamentous) 파지 발현, 분비 및/또는 디스플레이에 적합한 벡터를 포함한다. 소정의 예시적인 폴리뉴클레오티드가 본 명세서에 개시되지만, 주어진 발현 시스템에서의 유전 코드 또는 코돈 선호도의 축퇴를 고려했을 때, 본 발명의 단백질 스캐폴드 및 단백질 스캐폴드들의 라이브러리를 암호화하는 다른 폴리뉴클레오티드들이 또한 본 발명의 범주 내에 있다.
본 발명의 일 실시 형태는 서열 번호 18-29, 107-110, 또는 122-137의 아미노산 서열을 갖는, EGFR에 특이적으로 결합하는 FN3 도메인을 암호화하는 단리된 폴리뉴클레오티드이다.
본 발명의 일 실시 형태는 서열 번호 97-98 또는 168-169의 폴리뉴클레오티드 서열을 포함하는 단리된 폴리뉴클레오티드이다.
본 발명의 일 실시 형태는 서열 번호 32-49 또는 111-114에 나타낸 서열의 아미노산 서열을 갖는, c-Met에 특이적으로 결합하는 FN3 도메인을 암호화하는 단리된 폴리뉴클레오티드이다.
본 발명의 일 실시 형태는 서열 번호 50-72, 106, 118-121 또는 138-165의 아미노산 서열을 갖는 이중특이성 EGFR/-c-Met FN3 도메인 함유 분자를 암호화하는 단리된 폴리뉴클레오티드이다.
본 발명의 일 실시 형태는 서열 번호 115-116 또는 166-167의 폴리뉴클레오티드 서열을 포함하는 단리된 폴리뉴클레오티드이다.
본 발명의 폴리뉴클레오티드는 자동화 폴리뉴클레오티드 합성기 상에서 고상(solid phase) 폴리뉴클레오티드 합성과 같은 화학적 합성에 의해 제조되고 완전한 단일 또는 이중 가닥 분자로 조립될 수 있다. 대안적으로, 본 발명의 폴리뉴클레오티드는 PCR 후의 일상적인 클로닝과 같은 다른 기법에 의해 생성될 수 있다. 주어진 알려진 서열의 폴리뉴클레오티드를 생성하거나 얻는 기법은 당업계에 잘 알려져 있다.
본 발명의 폴리뉴클레오티드는 적어도 하나의 비암호화 서열, 예컨대 프로모터 또는 인핸서 서열, 인트론, 폴리아데닐화 신호, RepA 결합을 촉진시키는 cis 서열 등을 포함할 수 있다. 폴리뉴클레오티드 서열은 또한, 예를 들어, 마커 또는 태그 서열, 예컨대 단백질의 정제 또는 검출을 용이하게 하기 위한 히스티딘 태그 또는 HA 태그, 신호 서열, 융합 단백질 파트너, 예컨대 RepA, Fc 또는 박테리오파지 외피 단백질, 예컨대 pIX 또는 pIII을 암호화하는 추가 아미노산을 암호화하는 추가 서열을 포함할 수 있다.
본 발명의 다른 실시 형태는 본 발명의 적어도 하나의 폴리뉴클레오티드를 포함하는 벡터이다. 그러한 벡터는 플라스미드 벡터, 바이러스 벡터, 배큘로바이러스(baculovirus) 발현용 벡터, 트랜스포손(transposon) 기반 벡터, 또는 임의의 수단에 의해 주어진 유기체 또는 유전적 백그라운드(genetic background) 내로 본 발명의 폴리뉴클레오티드를 도입시키기에 적합한 임의의 다른 벡터일 수 있다. 그러한 벡터는 그러한 벡터에 의해 암호화되는 폴리펩티드의 발현을 제어하거나, 조절하거나, 야기하거나, 허용할 수 있는 핵산 서열 요소를 포함하는 발현 벡터일 수 있다. 그러한 요소는 전사 인핸서 결합 부위, RNA 폴리머라제 개시 부위, 리보솜 결합 부위, 및 주어진 발현 시스템에서 암호화된 폴리펩티드의 발현을 용이하게 하는 기타 부위를 포함할 수 있다. 그러한 발현 시스템은 당업계에 잘 알려진 세포 기반 시스템 또는 무세포 시스템일 수 있다.
본 발명의 다른 실시 형태는 본 발명의 벡터를 포함하는 숙주 세포이다. 본 발명의 단일특이성 EGFR-결합 또는 c-Met-결합 FN3 도메인 또는 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는, 당업계에 잘 알려진 바와 같이, 선택적으로 세포주, 혼합 세포주, 불멸화 세포 또는 불멸화 세포들의 클론 집단에 의해 생성될 수 있다. 예를 들어, 문헌[Ausubel, et al., ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, NY (1987-2001)]; 문헌[Sambrook, et al., Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor, NY (1989)]; 문헌[Harlow and Lane, Antibodies, a Laboratory Manual, Cold Spring Harbor, NY (1989)]; 문헌[Colligan, et al., eds., Current Protocols in Immunology, John Wiley & Sons, Inc., NY (1994-2001)]; 문헌[Colligan et al., Current Protocols in Protein Science, John Wiley & Sons, NY, NY, (1997-2001)]을 참조한다.
발현용으로 선택된 숙주 세포는 포유류 기원의 것일 수 있거나, 또는 COS-1, COS-7, HEK293, BHK21, CHO, BSC-1, He G2, SP2/0, HeLa, 골수종, 림프종, 효모, 곤충 또는 식물 세포, 또는 이들의 임의의 유도체, 불멸화 또는 형질전환 세포로부터 선택될 수 있다. 대안적으로, 숙주 세포는, 폴리펩티드를 글리코실화할 수 없는 종 또는 유기체, 예를 들어 원핵 세포 또는 유기체, 예컨대 BL21, BL21(DE3), BL21-GOLD(DE3), XL1-Blue, JM109, HMS174, HMS174(DE3), 및 천연 또는 조작된 E. 콜라이 종(E. coli spp.), 클렙시엘라 종(Klebsiella spp.), 또는 슈도모나스 종(Pseudomonas spp.) 균주 중 임의의 것으로부터 선택될 수 있다.
본 발명의 다른 실시 형태는 본 발명의 EGFR 또는 c-Met에 특이적으로 결합하는 단리된 FN3 도메인, 또는 본 발명의 단리된 이중특이성 EGFR/c-Met FN3 도메인 함유 분자를 생성하는 방법으로서, 본 방법은 EGFR 또는 c-Met에 특이적으로 결합하는 단리된 FN3 도메인 또는 단리된 이중특이성 EGFR-c-Met FN3 도메인 함유 분자가 발현되도록 하는 조건 하에서 본 발명의 단리된 숙주 세포를 배양하는 단계, 및 도메인 또는 분자를 정제하는 단계를 포함한다.
본 발명의 EGFR 또는 c-Met에 특이적으로 결합하는 FN3 도메인 또는 단리된 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 잘 알려진 방법에 의해, 예를 들어 단백질 A 정제, 황산암모늄 또는 에탄올 침전, 산 추출, 음이온 또는 양이온 교환 크로마토그래피, 포스포셀룰로스 크로마토그래피, 소수성 상호작용 크로마토그래피, 친화성 크로마토그래피, 하이드록실아파타이트 크로마토그래피 및 렉틴 크로마토그래피, 또는 고성능 액체 크로마토그래피(HPLC)에 의해 재조합 세포 배양물로부터 정제될 수 있다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자 및 EGFR-결합 또는 c-Met-결합 FN3 도메인의 용도
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인은 인간 질병 또는 세포, 조직, 기관, 체액, 또는 일반적으로 숙주에서의 특정 병상의 증상을 진단하거나, 모니터링하거나, 조절하거나, 치료하거나, 경감시키거나, 그의 발생의 방지를 돕거나, 또는 감소시키는 데 사용될 수 있다. 본 발명의 방법은 임의의 분류에 속하는 동물 환자를 치료하기 위해 사용될 수 있다. 그러한 동물의 예에는 포유동물, 예컨대 인간, 설치류, 개, 고양이 및 가축이 포함된다.
본 발명의 일 태양은 EGFR 및/또는 c-Met를 발현하는 세포의 성장 또는 증식을 억제하기 위한 방법으로서, 본 방법은 이러한 세포를 본 발명의 단리된 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인과 접촉시키는 단계를 포함한다.
본 발명의 다른 태양은 대상체에서 EGFR- 및/또는 c-Met-발현 종양 또는 암 세포의 성장 또는 전이를 억제하기 위한 방법으로서, 본 방법은 EGFR- 및/또는 c-Met-발현 종양 또는 암 세포의 성장 또는 전이가 억제되도록 유효량의 본 발명의 단리된 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인을 대상체에게 투여하는 단계를 포함한다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인은 EGFR, c-Met, EGF 또는 다른 EGFR 리간드 또는 HGF의 비정상 활성화 또는 생성을 특징으로 하는 임의의 질병 또는 장애, 또는 EGFR 또는 c-Met 발현과 관련된 장애 - 이러한 질병 또는 장애는 악성종양 또는 암을 포함할 수 있거나 그렇지 않을 수 있음 - 를 치료하는 데 사용될 수 있으며, 여기서 EGFR, c-Met, EGF 또는 다른 EGFR 리간드, 또는 HGF의 비정상 활성화 및/또는 생성은 이러한 질병 또는 장애를 갖거나 또는 이에 취약한 대상체의 세포 또는 조직 중에서 발생되고 있다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자는 암 및 양성 종양을 포함하는 종양의 치료에 사용될 수 있다. 본 발명의 이중특이성 분자에 의해 치료하기에 적합한 암은 EGFR 및/또는 c-Met를 과발현하는 것들을 포함한다. 본 발명의 이중특이성 분자에 의해 치료하기에 적합한 예시적인 암은 상피 세포암, 유방암, 난소암, 폐암, 비소세포 폐암(NSCLC), 폐 선암종, 결직장암, 항문암, 전립선암, 신장암, 방광암, 두경부암, 난소암, 췌장암, 피부암, 구강암, 식도암, 질암, 자궁경부암, 비장암, 고환암, 위암, 흉선암, 결장암, 갑상선암, 간암, 또는 산발성 또는 유전성 유두상 신장 암종(PRCC)을 포함한다.
c-Met에 특이적으로 결합하여 c-Met에 대한 HGF의 결합을 차단하는 본 발명의 FN3 도메인은 암 및 양성 종양을 포함하는 종양의 치료를 위한 것일 수 있다. 본 발명의 c-Met 결합 FN3 도메인에 의해 치료하기에 적합한 암은 c-Met를 과발현하는 것들을 포함한다. 본 발명의 FN3 도메인에 의해 치료하기에 적합한 예시적인 암은 상피 세포암, 유방암, 난소암, 폐암, 결직장암, 항문암, 전립선암, 신장암, 방광암, 두경부암, 난소암, 췌장암, 피부암, 구강암, 식도암, 질암, 자궁경부암, 비장암, 고환암, 및 흉선암을 포함한다.
EGFR에 특이적으로 결합하여 EGFR에 대한 EGF의 결합을 차단하는 본 발명의 FN3 도메인은 암 및 양성 종양을 포함하는 종양을 치료하는 데 사용될 수 있다. 본 발명의 FN3 도메인에 의해 치료하기에 적합한 암은 EGFR 또는 변이체를 과발현하는 것들을 포함한다. 본 발명의 FN3 도메인에 의해 치료하기에 적합한 예시적인 암은 상피 세포암, 유방암, 난소암, 폐암, 결직장암, 항문암, 전립선암, 신장암, 방광암, 두경부암, 난소암, 췌장암, 피부암, 구강암, 식도암, 질암, 자궁경부암, 비장암, 고환암, 및 흉선암을 포함한다.
투여/약제학적 조성물
치료적 용도를 위하여, 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR-결합 FN3 도메인 또는 c-Met-결합 FN3 도메인은 약제학적으로 허용되는 담체 중에 활성 성분으로서 유효량의 이러한 도메인 또는 분자를 함유하는 약제학적 조성물로서 제조될 수 있다. 용어 "담체"는 활성 화합물과 함께 투여되는 희석제, 애쥬번트(adjuvant), 부형제, 또는 비히클을 지칭한다. 이러한 비히클은 석유, 동물, 식물, 또는 합성 기원의 것들, 예컨대 낙화생유, 대두유, 광유, 참기름 등을 포함하는, 물 및 오일과 같은 액체일 수 있다. 예를 들어, 0.4% 염수 및 0.3% 글리신이 사용될 수 있다. 이들 용액은 무균성이고 일반적으로 미립자 물질이 없다. 이들은 종래의 잘 알려진 멸균 기법(예를 들어, 여과)에 의해 멸균될 수 있다. 조성물은 pH 조정제 및 완충제, 안정제, 증점제, 윤활제 및 착색제 등과 같은 생리적 조건에 근접시키기 위하여 필요한, 약제학적으로 허용되는 보조 물질을 함유할 수 있다. 그러한 약제학적 제형에서 본 발명의 분자의 농도는 광범위하게, 즉 약 0.5 중량% 미만, 통상 약 1 중량% 또는 적어도 약 1 중량%부터 많게는 15 또는 20 중량%까지 변동될 수 있으며, 선택된 특정 투여 방식에 따라, 필요 용량, 유체 부피, 점도 등에 기초하여 주로 선택될 것이다. 다른 인간 단백질, 예를 들어 인간 혈청 알부민을 포함하는 적합한 비히클 및 제형이, 예를 들어 문헌[Remington: The Science and Practice of Pharmacy, 21st Edition, Troy, D.B. ed., Lipincott Williams and Wilkins, Philadelphia, PA 2006, Part 5, Pharmaceutical Manufacturing pp 691-1092(특히 pp. 958-989 참조)]에 기재되어 있다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인의 치료적 용도를 위한 투여 방식은 이러한 작용제를 숙주에게 전달하는 임의의 적합한 경로일 수 있으며, 예컨대 비경구 투여, 예를 들어 진피내, 근육내, 복막내, 정맥내 또는 피하, 폐; 경점막(구강, 비강내, 질내, 직장) 투여로서, 이러한 투여에서는 정제, 캡슐, 용액, 분말, 겔, 입자 형태이고, 주사기, 이식 디바이스, 삼투압 펌프, 카트리지, 마이크로펌프 내에 담긴 제형; 또는 당업자에 의해 인식되는 다른 수단을 사용하는데, 이는 당업계에 잘 알려진 바와 같다. 부위 특이적 투여는, 예를 들어, 관절내, 기관지내, 복강내, 관절낭내, 연골내, 강내, 체강내, 소뇌내, 뇌실내, 결장내, 자궁경부내, 위내, 간내, 심근내, 골내, 골반내, 심장주위내, 복막내, 흉막내, 전립선내, 폐내, 직장내, 신장내, 망막내, 척수내, 활막내, 흉부내, 자궁내, 혈관내, 방광내, 병변내, 질, 직장, 협측, 설하, 비강내, 또는 경피 전달에 의해 달성될 수 있다.
따라서, 근육내 주사를 위한 본 발명의 약제학적 조성물은 1 ml의 멸균 완충수, 및 약 1 ng 내지 약 100 mg, 예를 들어 약 50 ng 내지 약 30 mg 또는 더 바람직하게는 약 5 mg 내지 약 25 mg의 본 발명의 FN3 도메인을 함유하도록 제조될 수 있다. 유사하게, 정맥내 주입을 위한 본 발명의 약제학적 조성물은 약 250 ml의 멸균 링거액, 및 약 1 mg 내지 약 30 mg, 예를 들어 약 5 mg 내지 약 25 mg의 본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR 결합 FN3 도메인 또는 c-Met 결합 FN3 도메인을 함유하도록 구성될 수 있다. 비경구로 투여가능한 조성물을 제조하기 위한 실제의 방법은 잘 알려져 있으며, 예를 들어 문헌["Remington's Pharmaceutical Science", 15th ed., Mack Publishing Company, Easton, PA]에 더욱 상세히 기재되어 있다.
본 발명의 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR-결합 FN3 도메인 또는 c-Met-결합 FN3 도메인은 저장을 위해 동결건조되고, 사용하기 전에 적합한 담체 중에서 재구성될 수 있다. 이 기법은 종래의 단백질 제제에 유효한 것으로 밝혀져 있으며, 당업계에 공지된 동결건조 및 재구성 기법이 사용될 수 있다.
이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR-결합 FN3 도메인 또는 c-Met-결합 FN3 도메인은 단회 용량으로 대상체에게 투여될 수 있거나, 또는, 예를 들어 1일, 2일, 3일, 5일, 6일, 1주, 2주, 3주, 1개월, 5주, 6주, 7주, 2개월 또는 3개월 후에 투여가 반복될 수 있다. 반복 투여는 동일한 용량으로 또는 상이한 용량으로 행해질 수 있다. 투여는 1회, 2회, 3회, 4회, 5회, 6회, 7회, 8회, 9회, 10회, 또는 그 이상 반복될 수 있다.
이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR-결합 FN3 도메인 또는 c-Met-결합 FN3 도메인은 동시적으로, 순차적으로 또는 개별적으로 제2 치료제와 병용하여 투여될 수 있다. 제2 치료제는 화학치료제, 항혈관생성제, 또는 세포독성 약물일 수 있다. 암을 치료하는 데 사용되는 경우, 이중특이성 EGFR/c-Met FN3 도메인 함유 분자, EGFR-결합 FN3 도메인 또는 c-Met-결합 FN3 도메인은 종래의 암 요법, 예컨대 수술, 방사요법, 화학요법 또는 이들의 조합과 병용하여 사용될 수 있다. 본 발명의 FN3 도메인과 병용하여 사용될 수 있는 예시적인 작용제는 HER2, HER3, HER4, VEGF의 길항제, 및 단백질 티로신 키나제 억제제, 예컨대 이레사®(게피티닙) 및 타르세바(에를로티닙)이다.
본 발명을 일반적인 개념으로 설명하였지만, 본 발명의 실시 형태는 하기 실시예에서 추가로 개시될 것이며, 이때 실시예는 청구범위의 범주를 제한하는 것으로 해석되어서는 안 된다.
실시예 1: 텐콘 라이브러리의 작제
텐콘(서열 번호 1)은 인간 테나신-C로부터의 15개의 FN3 도메인의 공통 서열로부터 설계된, 면역글로불린-유사 스캐폴드의 피브로넥틴 III형(FN3) 도메인이다(문헌[Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012]; 미국 특허 출원 공개제2010/0216708호). 텐콘의 결정 구조는 7개의 베타-가닥을 연결시키는 6개의 표면-노출된 루프를 보여준다. 이들 루프, 또는 각각의 루프 내의 선택된 잔기들은, 특정 표적에 결합하는 신규한 분자들을 선택하는 데 사용될 수 있는 피브로넥틴 III형(FN3) 도메인의 라이브러리를 작제하기 위하여 무작위화될 수 있다.
텐콘:
LPAPKNLVVSEVTEDSLRLSWTAPDAAFDSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVKGGHRSNPLSAEFTT(서열 번호 1):
TCL1 라이브러리의 작제
텐콘(서열 번호 1)의 FG 루프만을 무작위화하도록 설계된 라이브러리, TCL1을 cis-디스플레이 시스템과 함께 사용하기 위해 작제하였다(문헌[Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012]). 이 시스템에서는, Tac 프로모터를 위한 서열들, 텐콘 라이브러리 암호화 서열, RepA 암호화 서열, cis-요소, 및 ori 요소를 도입시킨 단일-가닥 DNA를 생성한다. 시험관내 전사/번역 시스템에서의 발현시에, 텐콘-RepA 융합 단백질의 복합체를 생성하는데, 이는 cis에서 그것을 암호화하는 DNA에 결합된다. 이어서, 표적 분자에 결합하는 복합체를 단리하고 폴리머라제 연쇄 반응(PCR)에 의해 증폭시키는데, 이는 하기에 기재된 바와 같다.
cis-디스플레이와 함께 사용하기 위한 TCL1 라이브러리의 작제를 PCR의 연속 라운드에 의해 2개의 절반 부분에서 최종 선형 이중 가닥 DNA 분자를 생성함으로써 달성하였으며; 5' 단편은 프로모터 및 텐콘 서열을 함유하며, 반면 3' 단편은 repA 유전자와 cis- 및 ori 요소를 함유한다. 전체 작제물을 생성하기 위하여 제한 효소분해(restriction digest)에 의해 이들 2개의 절반 부분을 조합한다. 텐콘의 FG 루프, KGGHRSN(서열 번호 86)에만 무작위 아미노산을 도입시키도록 TCL1 라이브러리를 설계하였다. 이 라이브러리의 작제에서는 NNS 코돈을 사용하였으며, 그 결과 FG 루프 내로의 모두 20개의 아미노산 및 하나의 정지 코돈의 도입이 가능하게 되었다. TCL1 라이브러리는 6개의 별개의 서브-라이브러리를 함유하며, 이들 각각은 다양성을 더욱 증가시키기 위하여 7 내지 12개 잔기의 상이한 무작위화된 FG 루프 길이를 갖는다. 텐콘-기반 라이브러리의 설계가 표 2에 나타나 있다.
[표 2]
TCL1 라이브러리를 작제하기 위하여, PCR의 연속 라운드를 수행하여 Tac 프로모터를 부착하고, F:G 루프 내로 축퇴를 구축하고, 최종 조립체를 위해 필요한 제한 부위들을 부가한다. 먼저, PCR에 의해 2개의 단계로 FG 루프의 프로모터 서열 및 텐콘 서열 5'을 함유하는 DNA 서열을 생성하였다. 완전 텐콘 유전자 서열에 상응하는 DNA를, 프라이머 POP2220(서열 번호 2) 및 TC5'toFG(서열 번호 3)를 이용하여 PCR 템플릿으로서 사용하였다. 이 반응으로부터 생성된 PCR 생성물을 프라이머 130mer(서열 번호 4) 및 Tc5'toFG를 이용하여 PCR 증폭의 다음 라운드를 위한 템플릿으로서 사용하여 텐콘에 대한 5' 및 프로모터 서열의 부착을 완료하였다. 다음으로, 순방향 프라이머 POP2222(서열 번호 5), 및 역방향 프라이머 TCF7(서열 번호 6), TCF8(서열 번호 7), TCF9(서열 번호 8), TCF10(서열 번호 9), TCF11(서열 번호 10), 또는 TCF12(서열 번호 11) - 이들은 축퇴 뉴클레오티드를 함유함 - 를 이용하여, 첫 번째 단계에서 생성된 DNA 생성물을 증폭시킴으로써 F:G 루프 내로 다양성을 도입시켰다. PCR 사이클을 최소화하고 라이브러리의 다양성을 최대화하기 위하여, 적어도 8회의 100 μL PCR 반응을 각각의 서브-라이브러리에 대해 수행하였다. 5 ㎍ 이상의 이 PCR 생성물을 겔 정제하고, 프라이머 POP2222(서열 번호 5) 및 POP2234(서열 번호 12)를 이용하여 후속 PCR 단계에서 사용하였으며, 그 결과 텐콘 서열의 3' 말단에 6xHis 태그 및 NotI 제한 부위가 부착되었다. 단지 15회의 PCR 사이클 및 500 ng 이상의 템플릿 DNA를 사용하여 이 PCR 반응을 수행하였다. 생성된 PCR 생성물을 겔 정제하고, NotI 제한 효소로 효소분해하고, 퀴아젠(Qiagen) 컬럼에 의해 정제하였다.
이 라이브러리의 3' 단편은 디스플레이를 위한 요소들을 함유하는 불변 DNA 서열이며, 이러한 요소들은 PspOMI 제한 부위, repA 유전자의 암호화 영역, 그리고 cis- 및 ori 요소를 포함한다. M13 순방향 및 M13 역방향 프라이머를 이용하여, 이 DNA 단편을 함유하는 플라스미드(pCR4Blunt)(인비트로젠(Invitrogen))를 사용하여 PCR 반응을 수행하였다. 생성된 PCR 생성물을 PspOMI에 의해 하룻밤 효소분해하고 겔 정제하였다. 라이브러리 DNA의 5' 부분을 repA 유전자를 함유하는 3' DNA에 연결하기 위하여, NotI 및 PspOMI 효소 및 T4 리가제의 존재 하에서 2 pmol의 5' DNA를 등몰량의 3' repA DNA에 연결하였다. 37℃에서 하룻밤 연결 후에, 연결된 DNA의 극히 일부분을 겔 상에서 이동시켜 연결 효율을 검사하였다. 연결된 라이브러리 생성물을 12회의 PCR 증폭으로 분할하였으며, 프라이머 쌍 POP2250(서열 번호 13)과 DidLigRev(서열 번호 14)를 이용하여 12-사이클 PCR 반응을 실시하였다. TCL1 라이브러리의 각각의 서브-라이브러리에 대한 DNA 산출량은 32 내지 34 ㎍의 범위였다.
이 라이브러리의 품질을 평가하기 위하여, 프라이머 Tcon5new2(서열 번호 15) 및 Tcon6(서열 번호 16)을 이용하여 작업 라이브러리의 극히 일부분을 증폭시켰으며, 리가제-비의존성 클로닝을 통해 변형된 pET 벡터 내로 클로닝하였다. 플라스미드 DNA를 BL21-GOLD(DE3) 컴피턴트 세포(competent cell)(스트라타진(Stratagene)) 내로 형질전환시켰으며, 96개의 무작위로 선택된 콜로니를 T7 프로모터 프라이머를 이용하여 서열화하였다. 어떠한 중복 서열도 발견되지 않았다. 전체적으로, 클론의 대략 70 내지 85%는 프레임-시프트 돌연변이 없이 완전한 프로모터 및 텐콘 암호화 서열을 가졌다. 정지 코돈을 갖는 클론을 제외한 기능적 서열 비율은 59% 내지 80%였다.
TCL2 라이브러리의 작제
텐콘의 BC 루프 및 FG 루프 둘 모두를 무작위화하고 각각의 위치에서의 아미노산들의 분포를 엄격하게 제어한 TCL2 라이브러리를 작제하였다. 표 3은 TCL2 라이브러리 내의 원하는 루프 위치들에서의 아미노산 분포를 나타낸다. 설계된 아미노산 분포는 2가지 목적을 가졌다. 첫째, 텐콘 결정 구조의 분석에 기초하여 그리고/또는 상동성 모델링으로부터 텐콘 접힘 및 안정성에 있어서 구조적으로 중요할 것으로 예측되는 잔기들 쪽으로 라이브러리를 편향시켰다. 예를 들어, 단지 소수성 아미노산들의 서브세트이도록 위치 29를 고정시켰는데, 이는 이 잔기를 텐콘 접힘부의 소수성 코어 내에 매립하였을 때이다. 설계의 제2 단계는, 고친화성 결합물질(high-affinity binder)을 효율적으로 생성하기 위해, 항체의 중쇄 HCDR3에서 우선적으로 발견되는 잔기들의 아미노산 분포 쪽으로 아미노산 분포를 편향시키는 것을 포함하였다(문헌[Birtalan et al., J Mol Biol 377:1518-28, 2008]; 문헌[Olson et al., Protein Sci 16:476-84, 2007]). 이러한 목적을 향하여, 표 3의 "설계된 분포"는 다음과 같은 분포를 지칭한다: 6% 알라닌, 6% 아르기닌, 3.9% 아스파라긴, 7.5% 아스파르트산, 2.5% 글루탐산, 1.5% 글루타민, 15% 글리신, 2.3% 히스티딘, 2.5% 아이소류신, 5% 류신, 1.5% 라이신, 2.5% 페닐알라닌, 4% 프롤린, 10% 세린, 4.5% 트레오닌, 4% 트립토판, 17.3% 티로신, 및 4% 발린. 이 분포에는 메티오닌, 시스테인, 및 정지 코돈이 없다.
[표 3]
TCL2 라이브러리의 5' 단편은 라이브러리 풀(슬로닝 바이오테크놀로지(Sloning Biotechnology))로서 화학적으로 합성된 텐콘(서열 번호 1)의 암호화 영역 및 프로모터를 함유하였다. 이 DNA 풀은 적어도 1 × 1011개의 상이한 구성원들을 함유하였다. 이 단편의 말단에, RepA에 대한 연결을 위한 설계시에 BsaI 제한 부위를 포함시켰다.
이 라이브러리의 3' 단편은 디스플레이를 위한 요소들을 함유하는 불변 DNA 서열이었으며, 이러한 요소들은 6xHis 태그, repA 유전자의 암호화 영역, 및 cis-요소를 포함한다. 기존의 DNA 템플릿(상기)과, 프라이머 LS1008(서열 번호 17) 및 DidLigRev(서열 번호 14)를 사용하여 PCR 반응에 의해 DNA를 제조하였다. 완전한 TCL2 라이브러리를 조립하기 위하여, 총 1 ㎍의 BsaI-효소분해된 5' 텐콘 라이브러리 DNA를, 동일한 효소를 이용하여 제한 효소분해에 의해 제조된 3.5 ㎍의 3' 단편에 연결하였다. 하룻밤 연결 후에, 퀴아젠 컬럼에 의해 DNA를 정제하였으며, 260 nm에서 흡광도를 측정함으로써 DNA를 정량화하였다. 연결된 라이브러리 생성물을 프라이머 쌍 POP2250(서열 번호 13)과 DidLigRev(서열 번호 14)를 이용하여 12-사이클 PCR 반응에 의해 증폭시켰다. 총 72회의 반응을 수행하였으며, 각각의 반응은 템플릿으로서 50 ng의 연결된 DNA 생성물을 포함하였다. TCL2 작업 라이브러리 DNA의 총 산출량은 약 100 ㎍이었다. 라이브러리 TCL1에 대해 전술된 바와 같이, 작업 라이브러리의 극히 일부분을 서브-클로닝하고 서열화하였다. 어떠한 중복 서열도 발견되지 않았다. 서열의 약 80%는 프레임-시프트 돌연변이 없이 완전한 프로모터 및 텐콘 암호화 서열을 함유하였다.
TCL14 라이브러리의 작제
상부(BC, DE, 및 FG) 및 하부(AB, CD, 및 EF) 루프, 예를 들어 FN3 도메인 내의 보고된 결합 표면은 FN3 구조의 중심을 형성하는 베타-가닥들에 의해 분리된다. 루프들만에 의해 형성된 표면과 상이한 형상들을 갖는 FN3 도메인의 2개의 "측(side)" 상에 존재하는 대체 표면은 2개의 역평행 베타-가닥, 즉 C 및 F 베타-가닥과, CD 및 FG 루프에 의해 FN3 도메인의 일측에 형성되고, 본 명세서에서 C-CD-F-FG 표면이라 칭한다.
도 4에 나타낸 바와 같이 C 및 F 가닥뿐만 아니라 CD 및 FG 루프의 부분들의 선택된 표면 노출된 잔기들을 무작위화함으로써 텐콘의 대체 표면을 무작위화하는 라이브러리를 생성하였다. 텐콘(서열 번호 1)과 비교할 때 하기의 치환을 갖는 텐콘27(서열 번호 99)의 텐콘 변이체를 사용하여 라이브러리를 생성하였다: E11R L17A, N46V, E86I. 이 라이브러리를 작제하는 데 사용되는 방법에 대한 충분한 설명이 미국 특허 출원 제13/852,930호에 기재되어 있다.
실시예 2: EGFR에 결합하여 EGF 결합을 억제하는 피브로넥틴 III형(FN3) 도메인의 선택
라이브러리 스크리닝
cis-디스플레이를 사용하여 TCL1 및 TCL2 라이브러리로부터 EGFR 결합 도메인을 선택하였다. IgG1 Fc에 융합된 EGFR의 재조합 인간 세포외 도메인(알앤디 시스템즈(R&D Systems))을 표준 방법을 사용하여 비오티닐화하고 패닝(panning)에 사용하였다(서열 번호 73의 전 길이 EGFR의 잔기 25-645). 시험관내 전사 및 번역(ITT)을 위해, 2 내지 6 ㎍의 라이브러리 DNA를, 100 μL의 총 부피로 0.1 mM의 완전 아미노산, 1X S30 프리믹스 성분들, 및 30 μL의 S30 추출물(프로메가(Promega))과 함께 30℃에서 인큐베이션하였다. 1시간 후에, 450 μL의 블로킹 용액(blocking solution)(2% 소혈청 알부민, 100 ㎍/mL의 청어 정자(herring sperm) DNA, 및 1 mg/mL의 헤파린으로 보충된 PBS pH 7.4)을 첨가하였으며, 반응물을 15분 동안 얼음 상에서 인큐베이션하였다. 실온에서 1시간 동안 블로킹 용액 중에서 재조합 인간 EGF(알앤디 시스템즈)를 비오티닐화 재조합 EGFR-Fc와 혼합함으로써 1:1 및 10:1의 EGFR 대 EGF의 몰비로 EGFR-Fc:EGF 복합체를 조립하였다. 결합을 위하여, 500 μL의 블로킹된 ITT 반응물을 100 μL의 EGFR-Fc:EGF 복합체와 혼합하고, 실온에서 1시간 동안 인큐베이션하였으며, 이후에 결합된 복합체를 뉴트라비딘 또는 스트렙타비딘 자성 비드(세라딘(Seradyn))를 사용하여 아래로 끌어당겼다. 결합되지 않은 라이브러리 구성원들을 PBST 및 PBS를 사용하여 연속 세척에 의해 제거하였다. 세척 후에, 10분 동안 65℃로 가열함으로써 결합된 복합체로부터 DNA를 용출하고, PCR에 의해 증폭시키고, 패닝의 추가 라운드를 위해 제한 효소분해 및 연결에 의해 RepA를 암호화하는 DNA 단편에 부착하였다. 각각의 라운드 동안 200 nM로부터 50 nM까지 표적 EGFR-Fc의 농도를 연속해서 낮추고 세척 엄격성(washing stringency)을 증가시킴으로써 고친화성 결합물질을 단리하였다.라운드 4 및 5에서는, PBS 중에서 하룻밤 10배 몰 과량의 비오티닐화되지 않은 EGFR-Fc의 존재 하에서 세척함으로써 결합되지 않은 FN3 도메인 및 약하게 결합된 FN3 도메인을 제거하였다.
패닝 후에, 선택된 FN3 도메인을 올리고(oligo) Tcon5new2(서열 번호 15) 및 Tcon6(서열 번호 16)을 사용하여 PCR에 의해 증폭시키고, 리가제 비의존성 클로닝 부위를 포함하도록 변형된 pET 벡터 내로 서브클로닝하고, 표준 분자 생물학 기법을 사용하여 E. 콜라이 내에서의 가용성 발현을 위하여 BL21-GOLD(DE3)(스트라타진) 세포 내로 형질전환시켰다. C-말단 폴리-히스티딘 태그를 암호화하는 유전자 서열을 각각의 FN3 도메인에 부가하여 정제 및 검출을 가능하게 하였다. 37℃에서 1 mL 96웰 블록들 내에서, 100 ㎍/mL의 카르베니실린으로 보충된 2YT 배지 중에서 배양물을 0.6 내지 0.8의 광학 밀도로 성장시켰으며, 그 후에 IPTG를 1 mM까지 첨가하였으며, 이 시점에서 온도를 30℃로 감소시켰다. 세포를 원심분리에 의해 대략 16시간 후에 수거하고, -20℃에서 동결시켰다. 45분 동안 실온에서 진탕하면서 0.6 mL의 버그버스터(BugBuster)® HT 용해 완충액(노바젠 이엠디 바이오사이언시스(Novagen EMD Biosciences)) 중에서 각각의 펠릿을 인큐베이션함으로써 세포 용해를 달성하였다.
세포 상에서의 EGFR에 결합하는 FN3 도메인의 선택
더 생리적인 환경에서 상이한 FN3 도메인들이 EGFR에 결합하는 능력을 평가하기 위하여, A431 세포에 결합하는 그들의 능력을 측정하였다. A431 세포(ATCC(American Type Culture Collection), 카탈로그 번호 CRL-1555)는 세포당 약 2 × 106개의 수용체로 EGFR을 과발현시킨다. 불투명한 흑색 96웰 플레이트들 내에서 세포를 5,000개/웰로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다.FN3 도메인-발현 세균 용해물(bacterial lysate)을 FACS 염색 완충액(벡톤 딕킨슨(Becton Dickinson)) 중에 1,000배로 희석시키고 3개의 반복된 플레이트 내에서 실온에서 1시간 동안 인큐베이션하였다. 용해물을 회수하였으며, 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였다. 실온에서 20분 동안 FACS 염색 완충액 중에 1:100으로 희석된 50 μL/웰의 항-펜타 히스-알렉사488(anti-penta His-Alexa488) 항체 접합체(퀴아젠)와 함께 세포를 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였으며, 이후에 웰들을 100 μL의 FACS 염색 완충액으로 충전하고, 아큐멘(Acumen) eX3 판독기를 사용하여 488 nm에서 형광을 판독하였다. FN3 도메인을 함유하는 세균 용해물을 A431 세포에 결합하는 그의 능력에 대해 스크리닝하였으며(TCL1 및 TCL2 라이브러리에 대해 1320개의 조(crude) 세포 용해물), 516개의 양성 클론을 확인하였는데, 여기서 결합은 백그라운드 신호에 비하여 10배 이상이었다. TCL14 라이브러리로부터의 300개의 용해물을 결합에 대해 스크리닝하였으며, 그 결과 58개의 특유의 FN3 도메인 서열이 EGFR에 대해 결합을 생성하였다.
세포 상에서의 EGFR에 대한 EGF의 결합을 억제하는 FN3 도메인의 선택
EGFR 결합의 기전을 보다 잘 특성화하기 위하여, 다양한 확인된 FN3 도메인 클론이 EGF-경쟁적 방식으로 EGFR에 결합하는 능력을 A431 세포를 사용하여 측정하였으며, A431 결합 검정 스크린과 병행하여 실시하였다. 불투명한 흑색 96웰 플레이트들 내에서 A431 세포를 5,000개/웰로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 3개의 반복된 플레이트 내에서 실온에서 1시간 동안 50 μL/웰의 1:1,000 희석된 세균 용해물과 함께 세포를 인큐베이션하였다. 비오티닐화 EGF(인비트로젠, 카탈로그 번호 E-3477)를 30 ng/mL의 최종 농도를 제공하도록 각각의 웰에 첨가하고, 실온에서 10분 동안 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였다. 실온에서 20분 동안 FACS 염색 완충액 중에 1:100으로 희석된 50 μL/웰의 스트렙타비딘-피코에리트린 접합체(인비트로젠)와 함께 세포를 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였으며, 이후에 웰들을 100 μL의 FACS 염색 완충액으로 충전하고, 아큐멘 eX3 판독기를 사용하여 600 nm에서 형광을 판독하였다.
FN3 도메인을 함유하는 세균 용해물을 전술된 EGF 경쟁 검정에서 스크리닝하였다. TCL1 및 TCL2 라이브러리로부터의 1320개의 조 세균 용해물을 스크리닝하였으며, 그 결과 451개의 양성 클론이 EGF 결합을 50% 초과로 억제하게 되었다.
EGFR에 결합하는 것으로 확인된 FN3 도메인의 발현 및 정제
히스 멀티트랩(His MultiTrap)™ HP 플레이트(지이 헬쓰케어(GE Healthcare))를 사용하여, 청징화된(clarified) E. 콜라이 용해물로부터 His-태깅된 FN3 도메인을 정제하였으며, pH 7.4에서 20 mM 인산나트륨, 500 mM 염화나트륨, 및 250 mM 이미다졸을 함유하는 완충액 중에서 용출시켰다. 정제된 샘플을 PD 멀티트랩™ G-25 플레이트(지이 헬쓰케어)를 사용하여 분석을 위하여 PBS pH 7.4 중으로 교환하였다.
크기 배제 크로마토그래피 분석
크기 배제 크로마토그래피를 사용하여 EGFR에 결합하는 FN3 도메인들의 응집 상태를 결정하였다. 각각의 정제된 FN3 도메인의 분취물들(10 μL)을 PBS pH 7.4의 이동상에서 0.3 mL/min의 유량으로 수퍼덱스(Superdex) 75 5/150 컬럼(지이 헬쓰케어) 상에 주입하였다. 컬럼으로부터의 용출을 280 nm에서의 흡광도에 의해 모니터링하였다. SEC에 의해 고수준의 응집을 나타낸 센티린(Centyrin)은 추가 분석으로부터 제외시켰다.
EGFR-Fc로부터의 선택된 EGFR-결합 FN3 도메인의 해리-속도(off-rate)
선택된 EGFR-결합 FN3 도메인을 스크리닝하여, 프로테온 XPR-36 기기(바이오라드)에서 EGFR-Fc에 결합시에 느린 해리-속도(koff)를 갖는 것들을 확인하여 고친화성 결합물질의 선택을 용이하게 하였다. 5 ㎍/mL 농도의 염소 항-인간 Fc IgG(알앤디 시스템즈)를, 칩 상에서 수평으로 배향된 모두 6개의 리간드 채널 상에서 0.005% 트윈(Tween)-20을 함유하는 PBS 중의 30 μL/min의 유량으로 (pH 5.0에서의) 아민 커플링을 통해 직접 고정화하였다. 고정화 밀도는 상이한 채널들 사이에 5% 미만의 변동을 갖고서 약 1500 반응 단위(Response Unit, RU)의 평균을 취하였다. 항-인간 Fc IgG 표면 상에 EGFR-Fc를 수직 리간드 배향으로 약 600 RU의 밀도로 포획하였다. 시험되는 모든 FN3 도메인을 1 μM의 농도에 대해 정규화하고 수평 배향으로 그들의 결합에 대해 시험하였다. FN3 도메인에 대해 모두 6개의 분석물 채널을 사용하여 스크리닝 처리량을 최대화하였다. 해리상(dissociation phase)을 100 μL/min의 유량으로 10분 동안 모니터링하였다. 분석물과 고정화된 IgG 표면 사이의 비특이적 결합을 모니터링하기 위해 스폿간(inter-spot) 결합 신호를 기준으로 사용하였으며, 이를 모든 결합 반응으로부터 제하였다. 처리된 결합 데이터를 1:1 단순 랭뮤어 결합 모델(simple Langmuir binding model)에 국소적으로 피팅하여, 포획된 EGFR-Fc에 대한 각각의 FN3 도메인의 결합에 대한 koff를 얻었다.
EGF-자극된 EGFR 인산화의 억제
정제된 EGFR-결합 FN3 도메인을 단일 농도로 A431 세포에서의 EGFR의 EGF-자극된 인산화를 억제하는 그의 능력에 대해 시험하였다. EGFR 포스포(Tyr1173) 키트(EGFR phospho(Tyr1173) kit)(메조 스케일 디스커버리(Meso Scale Discovery))를 사용하여 EGFR 인산화를 모니터링하였다. 투명한 96웰 조직 배양-처리된 플레이트들(눈크(Nunc)) 내에서, 10% 소태아 혈청(FBS)(깁코(Gibco))과 함께 글루타맥스(GlutaMAX)™를 함유하는 100 μL/웰의 RPMI 배지(깁코) 중에서 세포를 20,000개/웰로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 배양 배지를 완전히 제거하였으며, 가습된 5% CO2 분위기 내에서 37℃에서 FBS를 함유하지 않는 100 μL/웰의 배지 중에 세포를 하룻밤 기아상태에 두었다. 이어서, 가습된 5% CO2 분위기 내에서 37℃에서 1시간 동안 2 μM의 농도로 EGFR-결합 FN3 도메인을 함유하는 100 μL/웰의 예비가온된(37℃) 기아 배지(starvation medium)로 세포를 처리하였다. 대조군은 단지 기아 배지만으로 처리하였다. 100 ng/mL의 재조합 인간 EGF(알앤디 시스템즈, 카탈로그 번호 236-EG)를 함유하는 100 μL/웰의 예비가온된(37℃) 기아 배지를, 50 ng/mL의 EGF 및 1 μM의 EGFR-결합 FN3 도메인의 최종 농도가 되도록 첨가하고 온화하게 혼합하고, 15분 동안 37℃, 5% CO2에서 인큐베이션함으로써 세포를 자극하였다. 일 세트의 대조군 웰을 음성 대조군으로서 자극되지 않은 채로 그대로 두었다. 배지를 완전히 제거하였으며, 제조자의 사용설명서에 따라, 진탕하면서 실온에서 10분 동안 100 μL/웰의 완전 용해 완충액(Complete Lysis Buffer)(메조 스케일 디스커버리)으로 세포를 용해시켰다. 티로신 1173 상에서 인산화된 EGFR을 측정하도록 구성된 검정 플레이트(assay plate)(메조 스케일 디스커버리)를, 1.5 내지 2시간 동안 실온에서 제조자의 사용설명서에 따라, 제공된 블로킹 용액으로 블로킹하였다. 이어서, 플레이트를 200 μL/웰의 1X 트리스 세척 완충액(1X Tris Wash Buffer)(메조 스케일 디스커버리)으로 4회 세척하였다. 세포 용해물의 분취물들(30 μL/웰)을 검정 플레이트에 옮겼으며, 이를 플레이트 밀봉 필름(브이더블유알(VWR))으로 커버하고, 1시간 동안 진탕하면서 실온에서 인큐베이션하였다. 검정 플레이트를 200 μL/웰의 트리스 세척 완충액으로 4회 세척하였으며, 이후에 25 μL의 빙랭(ice-cold) 검출 항체 용액(Detection Antibody Solution)(메조 스케일 디스커버리)을 각각의 웰에 첨가하였는데, 이때 기포를 도입시키지 않도록 신중하게 하였다. 1시간 동안 진탕하면서 실온에서 플레이트를 인큐베이션한 후, 200 μL/웰의 트리스 세척 완충액으로 4회 세척하였다. 150 μL/웰의 판독 완충액(Read Buffer)(메조 스케일 디스커버리)을 첨가하고, 제조자-설정 검정-전용 디폴트 세팅(manufacturer-installed assay-specific default setting)을 사용하여 섹터(SECTOR)® 이미저 6000 기기(메조 스케일 디스커버리) 상에서 판독함으로써 신호를 검출하였다. EGF-자극된 양성 대조군 신호의 %억제율을 각각의 EGFR-결합 FN3 도메인에 대해 계산하였다.
EGF-자극된 EGFR 인산화의 억제를 TCL1 및 TCL2 라이브러리로부터의 232개의 확인된 클론에 대해 측정하였다. 이들 클론 중 22개는 1 μM 농도에서 EGFR 인산화를 50% 이상으로 억제하였다. 불량하게 발현되었거나 또는 크기 배제 크로마토그래피에 의해 다량체성인 것으로 판단된 클론을 제거한 후에, 9개의 클론을 추가의 생물학적 특성화를 위해 운반하였다. 이들 클론의 BC 및 FG 루프 서열이 표 4에 나타나 있다. 9개의 선택된 클론들 중 8개는 공통 FG 루프 서열(HNVYKDTNMRGL; 서열 번호 95)을 가졌으며, 상당한 유사성을 갖는 영역들이 몇몇 클론들 사이에서 그들의 BC 루프 서열에서 관찰되었다.
[표 4]
실시예 3: EGF 결합을 억제하는 EGFR-결합 FN3 도메인의 특성화
대규모 발현, 정제, 및 내독소 제거
표 4에 나타낸 9개의 FN3 도메인의 규모를 확대하여 상세한 특성화를 위한 더 많은 자료를 제공하였다. 각각의 EGFR-결합 FN3 도메인 변이체를 함유하는 하룻밤 배양물(overnight culture)을 사용하여, 하룻밤 배양물의 1/80 희석에서 100 ㎍/mL의 암피실린으로 보충된 0.8 L의 테리픽(Terrific) 브로쓰 배지를 새로운 배지 내로 접종하고, 37℃에서 진탕하면서 인큐베이션하였다. 600 nm에서의 광학 밀도가 약 1.2 내지 1.5에 도달했을 때 IPTG를 1 mM의 최종 농도가 되도록 첨가함으로써 배양을 유도하였으며, 온도를 30℃로 감소시켰다. 4시간 후에, 원심분리에 의해 세포를 수집하였으며, 필요할 때까지 세포 펠릿을 -80℃에서 저장하였다.
세포 용해를 위하여, 해동된 펠릿을 25 U/mL 벤조나제(Benzonase)®(시그마-알드리치(Sigma-Aldrich)) 및 1 kU/mL 알리소자임(rLysozyme)™(노바젠 이엠디 바이오사이언시스)으로 보충된 1X 버그버스터® 중에, 펠릿 1 g당 5 mL의 버그버스터®의 비로 재현탁시켰다. 온화하게 교반하면서 실온에서 1시간 동안 용해가 진행된 후, 4℃에서 50분 동안 56,000 × g로 원심분리를 행하였다. 상청액을 수집하고 0.2 μm 필터를 통해 여과하였으며, 이어서 지이 헬쓰케어 액타익스플로러(
) 100s 크로마토그래피 시스템을 사용하여 완충액 A(50 mM 트리스-HCl pH 7.5, 500 mM NaCl, 10 mM 이미다졸)에 의해 사전평형화된 5 mL 히스트랩(HisTrap) FF 컬럼 상에 로딩하였다. 컬럼을 20 컬럼 부피의 완충액 A로 세척하고, 6 컬럼 부피에 대한 16% 완충액 B(50 mM 트리스-HCl pH 7.5, 500 mM NaCl, 250 mM 이미다졸)로 추가로 세척하였다. FN3 도메인을 10 컬럼 부피에 대해 50% B로 용출한 후, 6 컬럼 부피에 걸쳐 50% B로부터 100% B까지의 구배를 행하였다. FN3 도메인 단백질을 함유하는 분획을 풀링하고, 밀리포어(Millipore) 10K MWCO 농축기를 사용하여 농축시키고, 여과한 후, PBS로 사전평형화된 하이로드(HiLoad)™ 16/60 수퍼덱스™ 75 컬럼(지이 헬쓰케어) 상에 로딩하였다. 크기 배제 컬럼으로부터 용출되는 단백질 단량체 피크를 보유하였다.
악티클린 에톡스(ActiClean Etox) 수지(스테로진 바이오세퍼레이션즈(Sterogene Bioseparations))를 이용하여 배치 접근(batch approach)을 사용하여 내독소를 제거하였다. 내독소 제거 전에, 37℃에서 2시간 동안(또는 4℃에서 하룻밤 동안) 1 N NaOH로 수지를 전처리하고, pH 시험지로 측정했을 때 pH가 약 7로 안정화될 때까지 PBS로 광범위하게 세척하였다. 정제된 단백질을 0.2 μm 필터를 통해 여과한 후, 1 mL의 에톡스 수지에 10 mL의 단백질 대 1 mL의 수지의 비로 첨가하였다. 수지에 대한 내독소의 결합은, 온화하게 회전시키면서 2시간 이상 동안 실온에서 진행되게 하였다. 2분 동안 500 × g로 원심분리에 의해 수지를 제거하였으며, 단백질 상청액을 보유하였다. 엔도세이프(EndoSafe)®-PTS™ 카트리지를 사용하여 내독소 수준을 측정하고, 엔도세이프®-MCS 판독기(찰스 리버(Charles River)) 상에서 분석하였다. 제1 에톡스 처리 후 내독소 수준이 5 EU/mg을 초과하였다면, 내독소 수준이 5 EU/mg 이하로 감소될 때까지 상기 절차를 반복하였다. 에톡스에 의한 2회의 연속 처리 후에 내독소 수준이 5 EU/mg을 초과하고 안정화된 경우에는, 단백질에 대해 음이온 교환 또는 소수성 상호작용 크로마토그래피 조건을 확립하여 남아 있는 내독소를 제거하였다.
EGFR-Fc에 대한 선택된 EGFR-결합 FN3 도메인의 친화도 결정(EGFR-Fc 친화도)
재조합 EGFR 세포외 도메인에 대한 선택된 EGFR-결합 FN3 도메인의 결합 친화도를 프로테온 기기(바이오라드)를 사용하여 표면 플라즈몬 공명 방법에 의해 추가로 특성화하였다. 검정 셋업(assay set-up)(칩 제조, EGFR-Fc 포획)은 해리-속도 분석에 대해 전술된 것과 유사하였다. 선택된 EGFR 결합 FN3 도메인을 수평 배향으로 3배 희석 시리즈로 1 μM 농도에서 시험하였다. 완충액 샘플을 또한 주입하여 기준선 안정성을 모니터링하였다. 각각의 EGFR-결합 FN3 도메인의 모든 농도에 대한 해리상을 30분 동안(해리-속도 스크리닝으로부터 koff가 약 10-2 s-1인 것들의 경우), 또는 1시간 동안(koff가 약 10-3 s-1 또는 그 미만인 것들의 경우) 100 μL/min의 유량으로 모니터링하였다. 2개의 세트의 기준 데이터를 반응 데이터로부터 제하였다: 1) 스폿간 신호, 이는 EGFR-결합 FN3 도메인과 고정화된 IgG 표면 사이의 비특이적 상호작용을 보정하기 위해서 제함; 2) 완충액 채널 신호, 이는 시간 경과에 따라 포획된 EGFR-Fc 표면의 해리로 인한 기준선 변화(baseline drifting)를 보정하기 위해서 제함. 각각의 FN3 도메인에 대한 모든 농도에서의 처리된 결합 데이터를 1:1 단순 랭뮤어 결합 모델에 전체적으로 피팅하여 속도론적 상수(kinetic constant)(kon, koff) 및 친화도 상수(K D)에 대한 추산치를 얻었다. 표 5는 작제물 각각에 대한 속도론적 상수를 나타내며, 이때 친화도는 200 pM부터 9.6 nM까지 다양하다.
세포 상에서의 EGFR에 대한 선택된 EGFR-결합 FN3 도메인의 결합(A431 세포 결합 검정)
불투명한 흑색 96웰 플레이트들 내에서 A431 세포를 5,000개/웰로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 정제된 EGFR-결합 FN3 도메인(1.5 nM 내지 30 μM)을 3개의 반복된 플레이트 내에서 실온에서 1시간 동안 (50 uL로) 세포에 첨가하였다. 상청액을 제거하였으며, 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였다. 실온에서 20분 동안 FACS 염색 완충액 중에 1:100으로 희석된 50 μL/웰의 항-펜타 히스-알렉사488 항체 접합체(퀴아젠)와 함께 세포를 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였으며, 이후에 웰들을 100 μL의 FACS 염색 완충액으로 충전하고, 아큐멘 eX3 판독기를 사용하여 488 nm에서 형광을 판독하였다. FN3 도메인 몰 농도의 로그에 대한 원시 형광 신호로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4(GraphPad Prism 4)(그래프패드 소프트웨어(GraphPad Software))를 사용하여 가변 기울기를 갖는 S자형 용량-반응 곡선에 피팅하여 EC50 값을 계산하였다. 표 5에는 작제물 각각에 대한 EC50이 2.2 내지 20 μM 초과의 범위인 것으로 기록되어 있다.
선택된 EGFR-결합 FN3 도메인을 사용하는, 세포 상에서의 EGFR에 대한 EGF 결합의 억제(A431 세포 EGF 경쟁 검정)
불투명한 흑색 96웰 플레이트들 내에서 A431 세포를 5,000개/웰로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 정제된 EGFR-결합 FN3 도메인(1.5 nM 내지 30 μM)을 3개의 반복된 플레이트 내에서 실온에서 1시간 동안 세포에 첨가하였다(50 μL/웰). 비오티닐화 EGF(인비트로젠, 카탈로그 번호 E-3477)를 30 ng/mL의 최종 농도를 제공하도록 각각의 웰에 첨가하고, 실온에서 10분 동안 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였다. 실온에서 20분 동안 FACS 염색 완충액 중에 1:100으로 희석된 50 μL/웰의 스트렙타비딘-피코에리트린 접합체(인비트로젠)와 함께 세포를 인큐베이션하였다. 세포를 150 μL/웰의 FACS 염색 완충액으로 3회 세척하였으며, 이후에 웰들을 100 μL의 FACS 염색 완충액으로 충전하고, 아큐멘 eX3 판독기를 사용하여 600 nm에서 형광을 판독하였다. FN3 도메인 몰 농도의 로그에 대한 원시 형광 신호로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4(그래프패드 소프트웨어)를 사용하여 가변 기울기를 갖는 S자형 용량-반응 곡선에 피팅하여 IC50 값을 계산하였다. 표 5에는 IC50 값이 1.8 내지 121 nM의 범위인 것으로 기록되어 있다.
EGF-자극된 EGFR 인산화의 억제(포스포-EGRF 검정)
EGF-자극된 EGFR 인산화를 유의하게 억제한 선택된 FN3 도메인을 억제에 대한 IC50 값을 측정함으로써 더 완벽히 평가하였다. "EGF 자극된 EGFR 인산화의 억제"에 전술된 바와 같이 FN3 도메인 농도들(0.5 nM 내지 10 μM)을 변동시킴으로써 EGF-자극된 EGFR 인산화의 억제를 평가하였다. FN3 도메인 몰 농도의 로그에 대한 전기화학발광 신호로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4(그래프패드 소프트웨어)를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 데이터를 피팅함으로써 IC50 값을 결정하였다. 표 5에는 IC50 값이 18 nM 내지 2.5 μM 초과의 범위인 것으로 기록되어 있다.
인간 종양 세포 성장의 억제(NCI-H292 성장 및 NCI-H322 성장 검정)
EGFR-결합 FN3 도메인에 대한 노출 후, EGFR 과발현 인간 종양 세포주, NCI-H292 및 NCI-H322(ATCC, 각각 카탈로그 번호 CRL-1848 및 CRL-5806)의 생존력을 측정함으로써 EGFR-의존성 세포 성장의 억제를 평가하였다. 불투명한 백색 96웰 조직 배양-처리된 플레이트들(눈크) 내에서, 10% 열 불활성화 소태아 혈청(깁코) 및 1% 페니실린/스트렙토마이신(깁코)으로 보충된 글루타맥스™ 및 10 mM HEPES를 함유하는 100 μL/웰의 RPMI 배지(깁코) 중에서 세포를 500 세포/웰(NCI-H292) 또는 1,000 세포/웰(NCI-H322)로 플레이팅하고, 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 일정 농도 범위의 EGFR-결합 FN3 도메인을 함유하는 5 μL/웰의 인산염 완충 식염수(PBS)를 첨가함으로써 세포를 처리하였다. 5 μL/웰의 PBS 단독 또는 PBS 중 25 mM 에틸렌다이아민테트라아세트산으로 대조군을 처리하였다. 120시간 동안 37℃, 5% CO2에서 세포를 인큐베이션하였다. 75 μL/웰의 셀타이터-글로(CellTiter-Glo)® 시약(프로메가)을 첨가한 후, 2분 동안 플레이트 진탕기(plate shaker) 상에서 혼합하고, 추가 10분 동안 실온에서 암소에서 인큐베이션함으로써 생존 세포(viable cell)를 검출하였다. 발광 모드로 설정된 스펙트라맥스(SpectraMax) M5 플레이트 판독기(몰레큘러 디바이시스(Molecular Devices)) 상에서 플레이트를 판독하였으며, 이때 판독 시간은 블랭크 상태의 배지 단독에 대해 0.5초/웰이었다. FN3 도메인 몰 농도의 로그에 대한 PBS-처리된 세포 성장의 백분율로서 데이터를 도표로 나타내었다. 그래프패드 프리즘 4(그래프패드 소프트웨어)를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 대한 식에 데이터를 피팅함으로써 IC50 값을 결정하였다. 표 5에는, NCI-H292 및 NCI-H322 세포를 사용했을 때,IC50 값이 각각 5.9 nM 내지 1.15 μM의 범위 및 9.2 nM 내지 3.1 μM 초과의 범위인 것으로 나타나 있다.
표 5에는 각각의 검정에 대한 EGFR-결합 FN3 도메인의 생물학적 특성에 대한 요약이 나타나 있다.
[표 5]
실시예 4: EGFR-결합 FN3 도메인의 조작
각각의 분자의 입체구조 안정성을 증가시키기 위해 EGFR 결합 FN3 도메인들의 서브세트를 조작하였다. 돌연변이 L17A, N46V 및 E86I(미국 특허 출원 공개 제2011/0274623호에 기재됨)를 DNA 합성에 의해 클론 P54AR4-83, P54CR4-31, 및 P54AR4-37 내로 도입시켰다. 전술된 바와 같이 새로운 돌연변이체 P54AR4-83v2, P54CR431-v2, 및 P54AR4-37v2를 발현시키고 정제하였다. PBS 중에서의 시차 주사 열량측정법을 사용하여 각각의 돌연변이체의 안정성을 평가하였는데, 이는 각각의 돌연변이체의 안정성을 상응하는 부모 분자의 안정성과 비교하기 위함이다. 표 6은 각각의 클론이 유의하게 안정화되었음을 보여주는데, 이때 Tm의 평균 증가는 18.5℃였다.
[표 6]
실시예 5: EGFR-결합 FN3 도메인의 시스테인 조작 및 화학적 접합
시스테인 잔기를 갖지 않는 베이스 텐콘 분자 및 그의 변이체로부터 FN3 도메인의 시스테인 돌연변이체를 제조한다. 이들 돌연변이는 소분자 약물, 형광 태그, 폴리에틸렌 글리콜, 또는 다수의 다른 화학적 실체의 화학적 접합을 위한 부위로서의 역할을 하기 위해 특유의 시스테인 잔기를 베이스 텐콘 서열(서열 번호 1) 또는 다른 FN3 도메인 내로 도입시키기 위해 당업계에 알려진 표준 분자 생물학 기법을 사용하여 행해질 수 있다. 선택하고자 하는 돌연변이의 부위는 소정의 기준을 만족해야 한다. 예를 들어, 유리 시스테인을 함유하도록 돌연변이화된 텐콘 분자는 (i) E. 콜라이(E. coli)에서 고도로 발현되어야 하고, (ii) 고수준의 용해도 및 열 안정성을 유지해야 하고, (iii) 접합 시에 표적 항원에 대한 결합을 유지해야 한다. 텐콘 스캐폴드는 단지 약 90 내지 95개의 아미노산으로 되어 있기 때문에, 단일-시스테인 변이체들이 스캐폴드의 매 위치마다 용이하게 작제될 수 있어서 화학적 접합에 대한 이상적인 위치(들)를 엄격히 결정할 수 있다.
EGFR과 결합하는 P54AR4-83v2 돌연변이체(서열 번호 27)의 위치 1 내지 95(또는 N-말단 메티오닌이 존재할 경우에는 2 내지 96)의 각각의 개별 아미노산 잔기를시스테인으로 돌연변이화하여 최상의 화학적 접합 부위를 평가하였다.
작제, 발현 및 정제
P54AR4-83v2의 각각의 개별 시스테인 변이체의 아미노산 서열을 E. 콜라이 발현을 위한 바람직한 코돈을 사용하여 단백질을 암호화하는 핵산 서열로 역변역시키고 합성 유전자를 생성하였다(DNA 2.0). 이들 유전자를 T5 프로모터 서열에 의해 구동되는 발현을 위한 pJexpress401 벡터(DNA 2.0) 내로 클로닝하고, E. 콜라이 균주 BL21(애질런트(Agilent)) 내로 형질전환시켰다. P54AR4-83v2 "cys 스캔" 라이브러리를 96웰 플레이트 내로 배열된 글리세롤 스톡으로서 제공하였으며, 각각의 발현 및 정제는 실시예 2에 기재된 것과 동일한 절차를 따랐다.
화학적 접합
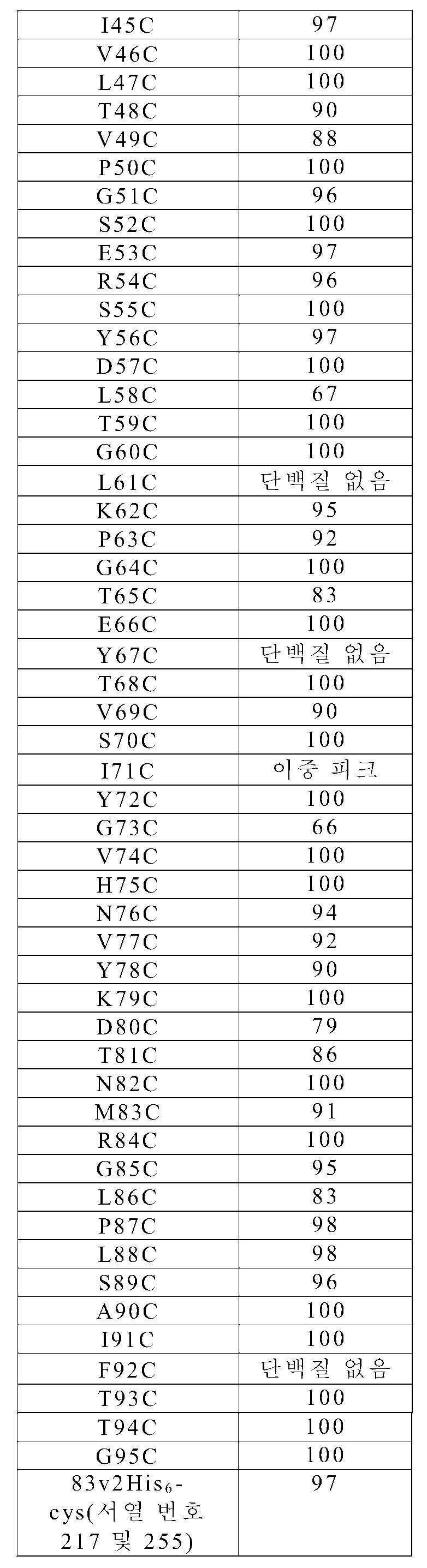
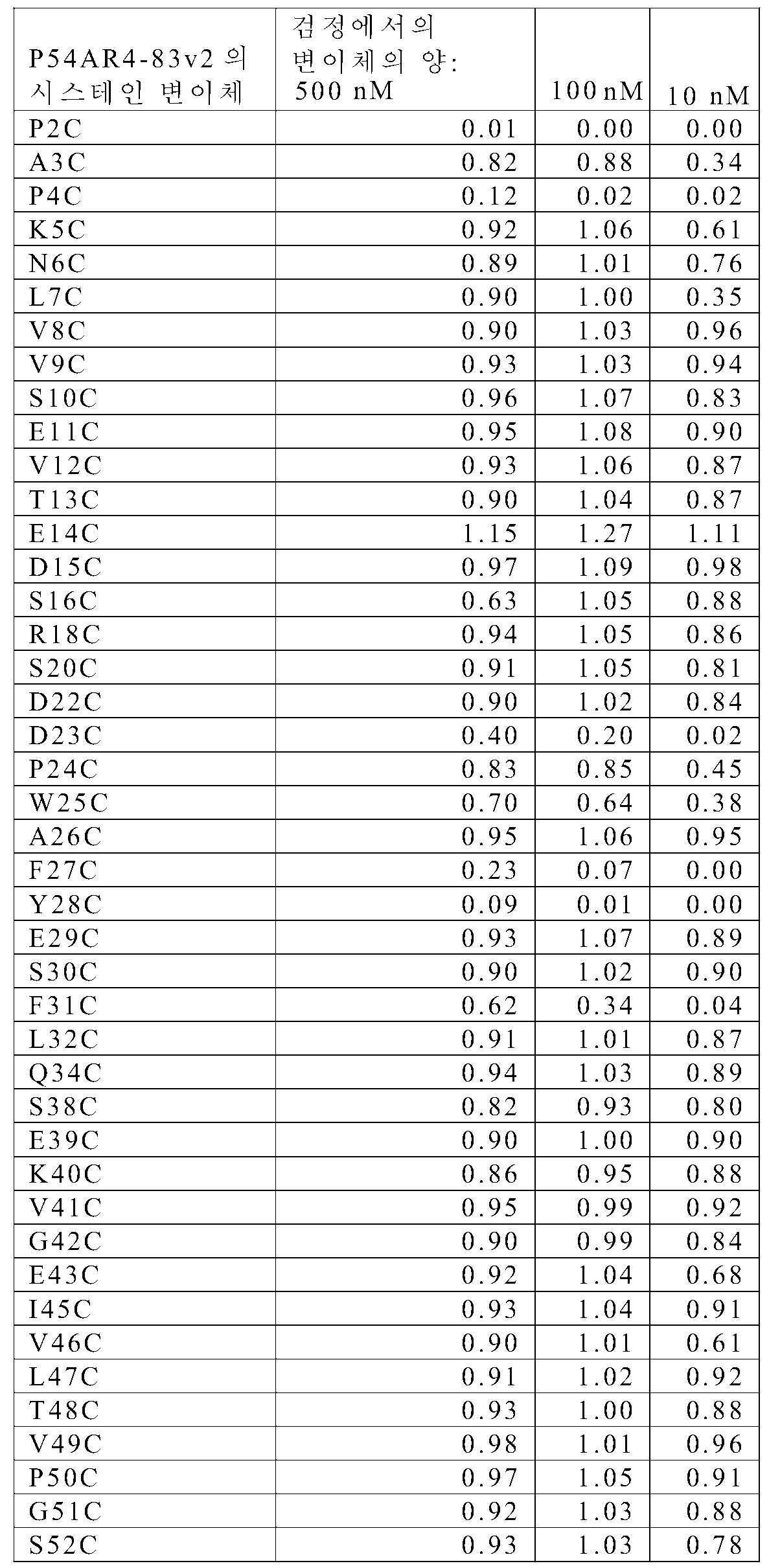
P54AR4-83v2 "cys 스캔" 라이브러리의 경우, 접합을 정제 공정 내로 통합시켰다. 청징화된 용해물 내의 시스테인 변이체를 히스-트랩 HP 플레이트(카탈로그 번호 28-4008-29, 지이 헬쓰케어)를 사용하여 96웰 포맷으로 Ni-NTA 수지에 결합하였는데, 이는 웰에 용해물을 첨가하고 5분 동안 100 × g로 원심분리함으로써 이루어졌다. 수지를 완충액 A로 3회 세척하고, 이어서 N-에틸 말레이미드(NEM)를 500 μL의 1.5 mM 용액으로서 첨가하였다. 로티세리 진탕기(rotisserie shaker) 상에서의 1시간의 실온 인큐베이션 후에, 원심분리하고 완충액 A로 3회 세척함으로써 과량의 NEM을 제거하였다. 접합된 시스테인 변이체를 2 × 150 μL의 완충액 B로 용출시키고, 울트라셀(Ultracel)-10 막을 갖는 멀티스크린 필터 플레이트(MultiScreen Filter Plate)(카탈로그 번호 MAUF1010, 밀리포어)를 사용하여 또는 96웰 PD 멀티트랩 플레이트(카탈로그 번호 28-9180-06, 지이 헬쓰케어)를 사용하여 PBS 중으로 교환하였다. 질량 분석에 의해 접합체를 특성화하였다(표 7). 불량하게 발현되거나(5 mL 배양액으로부터 얻어진 단백질이 0.1 mg 미만이거나 단백질이 질량 분석에 의해 검출되지 않거나) 또는 NEM에 불량하게 접합된(질량 분석에 의해 결정했을 때 80% 미만으로 접합된) 시스테인 변이체는 추가의 분석으로부터 제외시켰다. 이에 의해, 불량 발현으로 인해 L1C, W21C, Q36C, E37C, A44C, D57C, L61C, Y67C, 및 F92C를 그리고 낮은 접합 효율로 인해 A17C, L19C, I33C, Y35C, Y56C, L58C, T65C, V69C, I71C, 및 T94C를 제거하였다.
[표 7]
분석용 크기-배제 크로마토그래피
P54AR4-83v2의 각각의 NEM-접합된 시스테인 변이체에 대한 크기 배제 크로마토그래피를 실시예 2에 기재된 바와 같이 수행하였다. 표 8에 그 결과가 요약되어 있다. Abs280 신호를 통합시키고 단량체 영역(5.5 내지 6분) 내의 피크를 올리고머 영역 (4 내지 5.3분) 내의 피크들과 대비함으로써 각각의 단백질에 대한 %단량체를 결정하였다.
[표 8]
EGFR 결합 검정
EGFR에 대한 P54AR4-83v2의 NEM-접합된 시스테인 변이체들의 상대 결합 친화도를 실시예 2에 기재된 바와 같이 평가하였다. 표 9에는 P54AR4-83v2 부모 단백질에 대한 각각의 시스테인 변이체 EGFR 결합 친화도의 비를 보여주는 데이터가 요약되어 있다. ELISA 검정에 의해 결정했을 때 EGFR에 대해 감소된 결합을 가진(10 nM 단백질로 처리되었을 때 P54AR4-83v2 부모에 대해 관찰된 신호의 65% 미만인) 시스테인 접합체는 추가 분석으로부터 제외시켰다: P2C, A3C, P4C, K5C, L7C, D23C, W25C, F27C, Y28C, F31C, S55C, G73C, H75C, V77C, Y78C, T81C, N82C, M83C, 및 G85C.
[표 9]
열 안정성
시차 주사 열량계법(DSC)에 의해 시스테인-NEM 접합체의 열 안정성을 평가하였다. 단지 시험된 접합체들은 고수준으로 발현하는 것으로, 효율적으로 접합하는 것으로, 그리고 EGFR 결합을 유지하는 것으로 판단된 것들이었다. 추가적으로, BC 및 FG 루프 내의 시스테인 변이체는 제외시켰다. VP-DSC 기기(마이크로칼(MicroCal)) 내에서 분당 1℃의 스캔 속도로 25℃부터 100℃까지 변이체의 400 μL 분취물을 가열함으로써 안정성 데이터를 생성하였다. 열 접힘/풀림의 가역성을 평가하기 위해서 두 번째의 동일한 스캔을 샘플에 대해 완료하였다. 용융 온도를 계산하기 위하여 데이터를 2-상태 풀림 모델에 피팅하였다(표 10). 감소된 용융 온도를 갖거나(63℃ 이하이거나 P54AR4-83v2 부모보다 8℃ 초과하여 더 낮거나) 또는 비가역적 풀림을 나타낸 Cys 변이체는 추가의 분석으로부터 제외시켰다: V9C, V12C, T13C, R18C, E29C, E39C, G42C, V49C, P50C, G51C, P63C.
[표 10]
세포독성 검정
P54AR4-83v2시스테인 변이체를 NEM 접합에 대해 기재된 방법을 사용하여 세포독성 튜불린 억제제 모노메틸 오리스타틴 F(MMAF)에 효소-절단가능 Val-Cit 링커 또는 절단 불가능 PEG4 링커(VC-MMAF; 도 2 참조)를 통해 접합하였다.이전 단계에서 제외시킨 후에 남은 32개의 변이체를 P54AR4-83v2 부모(서열 번호217 및 255) 및 음성 대조군으로서의 텐콘(서열 번호 265)과 함께 접합시켰다.
EGFR-과발현 인간 종양 세포주 H1573을 시스테인 변이체-세포독소 접합체에 노출시킨 후에 그의 생존력을 측정함으로써 세포 사멸을 평가하였다. 흑색-웰의 투명 바닥의 조직 배양-처리된 플레이트들(팔콘(Falcon) 353219) 내에서, 5% 소태아 혈청(깁코)을 갖는 100 μL/웰의 페놀 레드 무함유 RPMI 배지(깁코 11835-030) 중에서 세포를 7000개/웰로 플레이팅하였다. 세포를 가습된 5% CO2 분위기 내에서 37℃에서 하룻밤 부착되게 하였다. 배지를 96웰 플레이트로부터 흡인하고, 세포를 50 uL의 새로운 배지 및 새로운 배지 중에 구성된 50 uL의 2X 억제제로 처리하였다. 70시간째에 셀타이터 글로(CellTiter Glo)(프로메가)를 사용하여 종점 검정에 의해 세포 생존력을 결정하였다. 그래프패드 프리즘 5(그래프패드 소프트웨어)를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 대한 식에 데이터를 피팅함으로써 IC50 값을 결정하였다. 표 11에는 셀타이터 글로 데이터의 분석으로부터 얻어진 IC50 값이 기록되어 있다. 83v2-cys/vcMMAF 접합체의 2회의 반복 실시의 평균 IC50은 0.7 nM이었다. 시험된 32개의 접합체 중 4개는 IC50 값이 부모의 것보다 2배 더 컸으며(1.4 nM 초과) 이들은 폐기하였다: L32C, T68C, Y72C, 및 V74C. 추가적으로, 3개의 접합체는 부모보다 2배 초과하여 더 강력한 IC50 값을 제공하였으며, 약물 접합체로 구성하는 데 특히 적합할 수 있다: N6C, E53C, 및 T93C.
[표 11]
최종 시스테인 변이체
시스테인 변이체들의 시험된 96개의 위치 중 28개의 위치가 E. 콜라이에서의 고발현 수준의 보유, 티올-말레이미드 화학적 반응을 통한 효율적인 접합, 표적 항원 EGFR에 대한 결합의 보유, 열 안정성 및 가역적 풀림 특성의 보유, 및 시스테인 변이체가 세포독성 약물에 접합될 때 고 EGFR 발현을 갖는 세포의 사멸의 보유의 기준을 만족시키는 것으로 확인되었다. 이들 위치는 다음과 같다: N6C(서열 번호 210 및 248), V8C(서열 번호 189 및 227), S10C(서열 번호 190 및 228), E11C(서열 번호 191 및 229), E14C(서열 번호 192 및 230), D15C(서열 번호 193 및 231), S16C(서열 번호 194 및 232), S20C(서열 번호 195 및 233), S30C(서열 번호 196 및 234), Q34C(서열 번호 197 및 235), S38C(서열 번호 198 및 236), K40C(서열 번호 199 및 237), V41C(서열 번호 200 및 238), I45C(서열 번호 201 및 239), L47C(서열 번호 202 및 240), T48C(서열 번호 203 및 241), E53C(서열 번호 204 및 242), R54C(서열 번호 205 및 243), T59C(서열 번호 206 및 244), G60C(서열 번호 207 및 245), K62C(서열 번호 208 및 246), G64C(서열 번호 209 및 247), T68C(서열 번호 210 및 248), S70C(서열 번호 211 및 249), L88C(서열 번호 212 및 250), S89C(서열 번호 213 및 251), A90C(서열 번호 214 및 252), I91C(서열 번호 215 및 253), 및 T93C(서열 번호 216 및 254). 83v2 단백질의 구조 내에서의 이들 28개의 위치들의 장소가 도 3에 나타나 있다.
실시예 6: c-Met에 결합하여 HGF 결합을 억제하는 피브로넥틴 III형(FN3) 도메인의 선택
인간 c-Met 상에서의 패닝
TCL14 라이브러리를 비오티닐화-인간 c-Met 세포외 도메인(bt-c-Met)에 대해 스크리닝하여 c-Met에 특이적으로 결합할 수 있는 FN3 도메인을 확인하였다. 선택을 위하여, E. 콜라이 S30 리니어 익스트랙트(Linear Extract)(미국 위스콘신주 메디슨 소재의 프로메가) 중에서 3 ㎍의 TCL14 라이브러리를 시험관내 전사 및 번역(IVTT)하였으며, 발현된 라이브러리를 시스 블록(Cis Block)(2% BSA(미국 미주리주 세인트루이스 소재의 시그마-알드리치), 100 ㎍/ml의 청어 정자 DNA(프로메가), 1 mg/mL의 헤파린(시그마-알드리치))으로 블로킹하였다. 선택을 위하여, bt-c-Met를 400 nM(라운드 1), 200 nM(라운드 2 및 3), 및 100 nM(라운드 4 및 5)의 농도로 첨가하였다. 뉴트라비딘 자성 비드(미국 일리노이주 록포드 소재의 써모 피셔(Thermo Fisher))(라운드 1, 3, 및 5) 또는 스트렙타비딘 자성 비드(프로메가)(라운드 2 및 4)를 사용하여 결합된 라이브러리 구성원을 회수하였으며, 비드를 500 uL의 PBS-T로 5 내지 14회 세척한 후 500 μL PBS로 2회 세척함으로써 결합되지 않은 라이브러리 구성원을 제거하였다.
개선된 친화도를 갖는 FN3 도메인 분자를 확인하기 위해 추가의 선택 라운드를 수행하였다. 간단히 말하면, 라운드 5로부터의 산출물을 전술된 바와 같이 제조하고, 하기의 변화를 동반하여 추가의 반복 선택 라운드를 거치게 하였다: bt-c-Met와의 인큐베이션을 1시간에서 15분으로 감소시켰고 비드 포획을 20분에서 15분으로 감소시켰으며, bt-c-Met를 25 nM(라운드 6 및 7) 또는 2.5 nM(라운드 8 및 9)로 감소시켰고, 과량의 비오티닐화되지 않은 c-Met의 존재 하에서 추가 1 시간의 세척을 수행하였다. 이들 변화의 목적은, 잠재적으로 더 빠른 결합 속도(on-rate) 및 더 느린 해리 속도를 가져서 실질적으로 더 낮은 K D를 산출하는 결합물질을 동시에 선택하기 위한 것이었다.
TCON6(서열 번호 30) 및 TCON5 E86I short(서열 번호 31) 프라이머를 이용하여 리가제 비의존성 클로닝 부위(pET15-LIC)를 함유하는 변형된 pET15 벡터(미국 뉴저지주 깁스타운 소재의 이엠디 바이오사이언시스) 내로 라운드 5, 7 및 9의 산출물을 PCR 클로닝하였으며, 이들 단백질은 표준 프로토콜을 사용하여 형질전환 및 IPTG 유도(최종 1 mM, 16시간 30℃)한 후에 C-말단 His6-태깅된 단백질로서 발현시켰다. 원심분리에 의해 세포를 수거하고, 이어서 0.2 mg/mL의 계난백 라이소자임(Chicken Egg White Lysozyme)(시그마-알드리치)으로 보충된 버그버스터 HT(이엠디 바이오사이언시스)로 용해시켰다. 세균 용해물을 원심분리에 의해 청징화하였으며, 상청액을 새로운 96 딥웰 플레이트에 옮겼다.
c-Met에 대한 HGF의 결합을 억제하는 FN3 도메인에 대한 스크리닝
E. 콜라이 용해물에 존재하는 FN3 도메인을 생화학적 포맷으로 정제된 c-Met 세포외 도메인에 대한 HGF 결합을 억제하는 그의 능력에 대해 스크리닝하였다. 재조합 인간 c-Met Fc 키메라(PBS 중 0.5 ㎍/mL, 100 μL/웰)를 96웰 백색 맥시소프 플레이트(White Maxisorp Plate)(눈크) 상에 코팅하고, 4℃에서 하룻밤 인큐베이션하였다. 바이오텍(Biotek) 플레이트 워셔 상에서 300 μl/웰의 0.05% 트윈 20을 함유하는 트리스 완충 식염수(TBS-T, 시그마-알드리치)로 플레이트를 2회 세척하였다. 진탕하면서 실온(RT)에서 1시간 동안 스타팅블록(StartingBlock) T20(200 μL/웰, 미국 일리노이주 록랜드 소재의 써모 피셔 사이언티픽(Thermo Fisher Scientific))을 사용하여 검정 플레이트를 블로킹하였으며, 300 μl의 TBS-T로 다시 2회 세척하였다. 해밀턴 스타 플러스(Hamilton STAR plus) 로보틱스 시스템을 사용하여, FN3 도메인 용해물을 스타팅블록 T20 중에 (1:10으로부터 1:100,000까지) 희석시켰다. 용해물(50 μL/웰)을 진탕하면서 실온에서 1시간 동안 검정 플레이트 상에서 인큐베이션하였다. 플레이트를 세척하지 않고서, bt-HGF(스타팅블록 T20 중 1 ㎍/mL, 50 μL/웰, 비오티닐화)를 진탕하면서 실온에서 30분 동안 플레이트에 첨가하였다. 텐콘27 용해물을 함유하는 대조군 웰은 스타팅 블록 T20 또는 희석된 bt-HGF를 수용하였다. 이어서, 플레이트를 300 μl/웰의 TBS-T로 4회 세척하고, 진탕하면서 실온에서 30 내지 40분 동안 100 μl/웰의 스트렙타비딘-HRP(TBS-T 중 1:2000, 미국 펜실베이니아주 웨스트 그로브 소재의 잭슨 이뮤노리서치(Jackson Immunoresearch))와 함께 인큐베이션하였다. 플레이트를 TBS-T로 다시 4회 세척하였다. 신호를 발생시키기 위하여, 제조자의 사용설명서에 따라 제조된 POD 화학발광 기질(POD Chemiluminescence Substrate)(50 μL/웰, 미국 인디애나주 인디애나폴리스 소재의 로슈 다이아그노스틱스(Roche Diagnostics))을 플레이트에 첨가하였으며, 대략 3분 이내에 소프트맥스 프로(SoftMax Pro)를 사용하여 몰레큘러 디바이시스 M5 상에서 발광을 판독하였다. 하기 계산을 사용하여 %억제율을 결정하였다: 100-((RLU샘플 - 평균 RLU무 bt-HGF 대조군)/(평균 RLUbt-HGF 대조군 - 평균 RLU무 bt-HGF 대조군)*100). 50% 이상의 %억제율 값을 히트(hit)로 간주하였다.
FN3 도메인의 대용량(high-throughput) 발현 및 정제
히스 멀티트랩™ HP 플레이트(지이 헬쓰케어)를 사용하여, 청징화된 E. 콜라이 용해물로부터 His-태깅된 FN3 도메인을 정제하였으며, pH 7.4에서 20 mM 인산나트륨, 500 mM 염화나트륨, 및 250 mM 이미다졸을 함유하는 완충액 중에서 용출시켰다. 정제된 샘플을 PD 멀티트랩™ G-25 플레이트(지이 헬쓰케어)를 사용하여 분석을 위하여 PBS pH 7.4 중으로 교환하였다.
c-Met에 대한 HGF 결합의 억제에 대한 IC
50
결정
선택된 FN3 도메인을 HGF 경쟁 검정에서 추가로 특성화하였다. 정제된 FN3 도메인에 대한 용량 반응 곡선을 전술된 검정을 이용하여 생성하였다(5 μM의 출발 농도). %억제율 값을 계산하였다. FN3 도메인 몰 농도의 로그에 대한 %억제율로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 데이터를 피팅함으로써 IC50 값을 결정하였다.
35개의 특유의 서열이 라운드 5로부터 1:10의 희석에서 활성을 나타내는 것으로 확인되었으며, 이때 IC50 값은 0.5 내지 1500 nM의 범위였다. 라운드 7은 39개의 특유의 서열을 산출하였는데, 이때 1:100의 희석에서 활성을 가졌으며, IC50 값은 0.16 내지 2.9 nM의 범위였다. 라운드 9로부터 66개의 특유의 서열을 확인하였는데, 여기서는 1:1000의 희석에서 활성인 것으로 히트를 규정하였다. 0.2 nM만큼이나 낮은 IC50 값이 라운드 9에서 관찰되었다(표 13).
실시예 7: c-Met에 결합하여 HGF 결합을 억제하는 FN3 도메인의 특성화
실시예 2에 전술된 바와 같이 FN3 도메인을 발현시키고 정제하였다. 실시예 1 및 실시예 2에 각각 전술된 바와 같이, 크기 배제 크로마토그래피 및 속도론적 분석을 행하였다. 표 12에는 각각의 도메인에 대한 C-가닥, CD 루프, F-가닥, 및 FG 루프의 서열, 및 전체 아미노산 서열에 대한 서열 번호가 나타나 있다.
[표 12]
C 루프 잔기는 표시된 서열 번호의 잔기 28-37에 상응한다.
CD 가닥 잔기는 표시된 서열 번호의 잔기 38-43에 상응한다.
F 루프 잔기는 표시된 서열 번호의 잔기 65-74에 상응한다.
FG 가닥 잔기는 표시된 서열 번호의 잔기 75-81에 상응한다.
세포 상에서의 c-Met에 대한 선택된 c-Met-결합 FN3 도메인의 결합
폴리-D-라이신 코팅된 흑색 투명 바닥 96웰 플레이트들(미국 캘리포니아주 새너제이 소재의 비디 바이오사이언시스(BD Biosciences)) 내에서 NCI-H441 세포(카탈로그 번호 HTB-174, 미국 버지니아주 매나사스 소재의 ATCC)를 20,000 세포/웰로 플레이팅하고, 37℃, 5% CO2에서 하룻밤 부착되게 하였다. 정제된 FN3 도메인(50 μL/웰; 0 내지 1000 nM)을 2개의 반복된 플레이트 내에서 4℃에서 1시간 동안 세포에 첨가하였다. 상청액을 제거하였으며, 세포를 FACS 염색 완충액(150 μL/웰, 비디 바이오사이언시스, 카탈로그 번호 554657)으로 3회 세척하였다. 4℃에서 30분 동안 비오티닐화-항 HIS 항체(FACS 염색 완충액 중에 1:160으로 희석됨, 50 μL/웰, 알앤디 시스템즈, 카탈로그 번호 BAM050)와 함께 세포를 배양하였다. 세포를 FACS 염색 완충액(150 μL/웰)으로 3회 세척하였으며, 이후에 4℃에서 30분 동안 항 마우스 IgG1-알렉사 488 접합된 항체(FACS 염색 완충액 중에 1:80으로 희석됨, 50 μL/웰, 라이프 테크놀로지스(Life Technologies), 카탈로그 번호 A21121)와 함께 웰을 인큐베이션하였다. 세포를 FACS 염색 완충액(150 μL/웰)으로 3회 세척하고, FACS 염색 완충액(50 μL/웰) 중에 그대로 두었다. 아큐멘 eX3 판독기를 사용하여 총 형광을 결정하였다. FN3 도메인 몰 농도의 로그에 대한 원시 형광 신호로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4(그래프패드 소프트웨어)를 사용하여 가변 기울기를 갖는 S자형 용량-반응 곡선에 피팅하여 EC50 값을 계산하였다. FN3 도메인은 일정 범위의 결합 활성을 나타내는 것으로 확인되었으며, 이때 EC50 값은 1.4 내지 22.0으로서, 이는 표 13에 나타낸 바와 같다.
HGF-자극된 c-Met 인산화의 억제
메조 스케일 디스커버리(미국 매릴랜드주 게이더스버그 소재)로부터의 c-Met 포스포(Tyr1349) 키트를 사용하여, 정제된 FN3 도메인을 NCI-H441에서의 c-Met의 HGF-자극된 인산화를 억제하는 그의 능력에 대해 시험하였다. 투명한 96웰 조직 배양-처리된 플레이트들 내에서, 10% 소태아 혈청(FBS; 라이프 테크놀로지스)과 함께 (글루타맥스 및 HEPES(라이프 테크놀로지스)를 함유하는) 100 μL/웰의 RPMI 배지 중에서 세포를 20,000개/웰로 플레이팅하고, 37℃, 5% CO2에서 하룻밤 부착되게 하였다. 배양 배지를 완전히 제거하였으며, 37℃, 5% CO2에서 무혈청 RPMI 배지(100 μL/웰) 중에 세포를 하룻밤 기아상태에 두었다. 이어서, 37℃, 5% CO2에서 1시간 미만 동안 그리고 20 μM 농도로 FN3 도메인을 함유하는 새로운 무혈청 RPMI 배지(100 μL/웰)를 세포에 보급하였다. 대조군은 배지 단독으로 처리하였다. 100 ng/mL의 재조합 인간 HGF(100 μL/웰, 알앤디 시스템즈, 카탈로그 번호 294-HGN)로 세포를 자극하고, 15분 동안 37℃, 5% CO2에서 인큐베이션하였다. 일 세트의 대조군 웰을 음성 대조군으로서 자극되지 않은 채로 그대로 두었다. 이어서, 배지를 완전히 제거하였으며, 제조자의 사용설명서에 따라, 진탕하면서 실온에서 10분 동안 완전 용해 완충액(50 μL/웰, 메조 스케일 디스커버리)으로 세포를 용해시켰다. 인산화된 c-Met를 측정하도록 구성된 검정 플레이트를, 1시간 동안 실온에서 제조자의 사용설명서에 따라, 제공된 블로킹 용액으로 블로킹하였다. 이어서, 플레이트를 트리스 세척 완충액(200 μL/웰, 메조 스케일 디스커버리)으로 3회 세척하였다. 세포 용해물(30 μL/웰)을 검정 플레이트에 옮기고, 1시간 동안 진탕하면서 실온에서 인큐베이션하였다. 이어서, 검정 플레이트를 트리스 세척 완충액으로 4회 세척하였으며, 이후에 빙랭 검출 항체 용액(25 μL/웰, 메조 스케일 디스커버리)을 진탕하면서 실온에서 1시간 동안 각각의 웰에 첨가하였다. 플레이트를 트리스 세척 완충액으로 다시 4회 헹구었다. 150 판독 완충액(150 μL/웰, 메조 스케일 디스커버리)을 첨가하고, 제조자-설정 검정-전용 디폴트 세팅을 사용하여 섹터® 이미저 6000 기기(메조 스케일 디스커버리) 상에서 판독함으로써 신호를 검출하였다. FN3 도메인 몰 농도의 로그에 대한 전기화학발광 신호로서 데이터를 도표로 나타내었으며, 그래프패드 프리즘 4를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 데이터를 피팅함으로써 IC50 값을 결정하였다. FN3 도메인은 인산화된 c-Met를 억제하는 것으로 확인되었는데, 이때 IC50 값은 4.6 내지 1415 nM의 범위로서, 이는 표 13에 나타낸 바와 같다.
인간 종양 세포 성장의 억제
c-Met-결합 FN3 도메인에 노출 후, U87-MG 세포(ATCC, 카탈로그 번호 HTB-14)의 생존력을 측정함으로써 c-Met-의존성 세포 성장의 억제를 평가하였다. 불투명한 백색 96웰 조직 배양-처리된 플레이트들(눈크) 내에서, 10% FBS로 보충된 100 μL/웰의 RPMI 배지 중에서 세포를 8000 세포/웰로 플레이팅하고, 37℃, 5% CO2에서 하룻밤 부착되게 하였다. 플레이팅 후 24시간째에, 배지를 흡인하였으며 세포에 무혈청 RPMI 배지를 보급하였다. 혈청 기아 후 24시간째에, c-Met-결합 FN3 도메인을 함유하는 무혈청 배지(30 μL/웰)를 첨가함으로써 세포를 처리하였다. 72시간 동안 37℃, 5% CO2에서 세포를 인큐베이션하였다. 100 μL/웰의 셀타이터-글로® 시약(프로메가)을 첨가한 후, 10분 동안 플레이트 진탕기 상에서 혼합함으로써 생존 세포를 검출하였다. 발광 모드로 설정된 스펙트라맥스 M5 플레이트 판독기(몰레큘러 디바이시스) 상에서 플레이트를 판독하였으며, 이때 판독 시간은 0.5초/웰이었다. FN3 도메인 몰 농도의 로그에 대한 미처리 발광 단위(raw luminescence unit, RLU)로서 데이터를 도표로 나타내었다. 그래프패드 프리즘 4를 사용하여 가변 기울기를 갖는 S자형 용량 반응에 대한 식에 데이터를 피팅함으로써 IC50 값을 결정하였다. 표 13에는 IC50 값이 1 nM 내지 1000 nM 초과의 범위인 것으로 기록되어 있다.
[표 13]
c-Met-결합 FN3 도메인의 열 안정성
PBS 중에서의 시차 주사 열량측정법을 사용하여 각각의 FN3 도메인의 안정성을 평가하였다. 실험 결과가 표 14에 나타나 있다.
[표 14]
실시예 8. 이중특이성 항-EGFR/c-Met 분자의 생성 및 특성화
이중특이성 EGFR/c-Met 분자의 생성
실시예 1 내지 실시예 6에 기재된 EGFR-결합 FN3 도메인과 c-Met-결합 FN3 도메인의 다수의 조합들을 연결시켜 EGFR 및 c-Met 둘 모두에 결합할 수 있는 이중특이성 분자로 하였다. 추가적으로, 서열 번호 107-110에 나타낸 아미노산 서열을 갖는 EGFR-결합 FN3 도메인 및 서열 번호 111-114에 나타낸 아미노산 서열을 갖는 c-Met 결합 FN3 도메인을 제조하고, 이들을 연결시켜 이중특이성 분자로 하였다. 하기 포맷, 즉 EGFR-결합 FN3 도메인, 그에 이어지는 펩티드 링커, 그에 이어지는 c-Met-결합 FN3 도메인을 유지하도록 서열 번호 50-72 및 106(표 15)에 기재된 아미노산 서열을 암호화하기 위해 합성 유전자를 생성하였다. 정제를 돕기 위해 폴리-히스티딘 태그를 C-말단에 도입시켰다.표 15에 기재된 분자들에 더하여, 이들 2개의 FN3 도메인들 사이의 링커는 표 16에 열거된 것들에 따라 길이, 서열 조성 및 구조에 따라 다양하였다. 다수의 다른 링커들이 그러한 FN3 도메인들을 연결하는 데 사용될 수 있었을 것으로 고려된다. IMAC 및 겔 여과 크로마토그래피 단계들을 사용하여 단일특이성 EGFR 또는 c-Met FN3 도메인에 대해 기재된 바와 같이 E. 콜라이로부터 이중특이성 EGFR/c-Met 분자를 발현시키고 정제하였다.
[표 15]
[표 16]
이중특이성 EGFR/c-Met 분자는 단일특이성 분자 단독과 비교하여 효력을 향상시킨다 - 결합력을 시사함
96웰 플레이트 내에서, 10% FBS를 함유하는 RPMI 배지 중에서 NCI-H292 세포를 플레이팅하였다. 24시간 후에, 배지를 무혈청 RPMI로 대체하였다. 혈청 기아 후 24시간째에, 하기 FN3 도메인들의 다양한 농도들로 세포를 처리하였다: 고 친화도 단일특이성 EGFR FN3 도메인(P54AR4-83v2), 약 친화도 단일특이성 c-Met FN3 도메인(P114AR5P74-A5), 2개의 단일특이성 EGFR과 c-Met FN3 도메인의 혼합물, 또는 고 친화도 EGFR FN3 도메인(ECB1)에 연결된 저 친화도 c-Met FN3 도메인으로 구성된 이중특이성 EGFR/c-Met 분자. 세포를 단일특이성 또는 이중특이성 분자로 1시간 동안 처리하고, 이어서 37℃, 5% CO2에서 15분 동안 EGF, HGF, 또는 EGF와 HGF의 조합으로 자극하였다. 세포를 MSD 용해 완충액으로 용해시켰으며, 전술된 바와 같이 제조자의 사용설명서에 따라 적절한 MSD 검정 플레이트를 사용하여 세포 신호전달을 평가하였다.
저 친화도 c-Met FN3 도메인은 610 nM의 IC50으로 c-Met의 인산화를 억제하였다(도 6). 예측된 바와 같이, EGFR FN3 도메인은 c-Met 인산화를 억제할 수 없었으며, 단일특이성 분자들의 혼합물은 c-Met FN3 도메인 단독과 동일한 것으로 보였다. 그러나, 이중특이성 EGFR/c-Met 분자는 1 nM의 IC50으로 c-Met의 인산화를 억제하여(도 6), c-Met 단일특이성 단독에 비하여 효력을 개선함에 있어서 2-로그 시프트를 초과하여 제공하였다.
이중특이성 EGFR/c-Met 분자가 결합력 효과를 통해 c-Met 및/또는 EGFR 인산화의 억제를 향상시킬 수 있는 잠재성을 c-Met 및 EGFR의 가변적인 밀도 및 비로 다수의 세포 유형에서 평가하였다(도 7). 96웰 플레이트 내에서, 10% FBS를 함유하는 RPMI 배지 중에서 NCI-H292, NCI-H441, 또는 NCI-H596 세포를 플레이팅하였다. 24시간 후에, 배지를 무혈청 RPMI로 대체하였다. 혈청 기아 후 24시간째에, 단일특이성 EGFR-결합 FN3 도메인, 단일특이성 c-Met FN3 도메인, 또는 이중특이성 EGFR/c-Met 분자(ECB5, 이는 P53A1R5-17v2 및 P114AR7P94-A3으로 구성됨)의 다양한 농도들로 세포를 처리하였다. 세포를 단일특이성 또는 이중특이성 분자로 1시간 동안 처리하고, 이어서 37℃, 5% CO2에서 15분 동안 EGF, HGF, 또는 EGF와 HGF의 조합으로 자극하였다. 세포를 MSD 용해 완충액으로 용해시켰으며, 전술된 바와 같이 제조자의 사용설명서에 따라 적절한 MSD 검정 플레이트를 사용하여 세포 신호전달을 평가하였다.
도 7(A 내지 C)은 3개의 상이한 세포주에서 이중특이성 EGFR/cMet 분자와 비교하여 단일특이성 EGFR-결합 FN3 도메인을 사용하여 EGFR을 억제하는 것을 나타낸다. EGFR 인산화 검정에서 결합력을 평가하기 위하여, 중간 친화도 EGFR-결합 FN3 도메인(1.9 nM)(P53A1R5-17v2)을, 고 친화도 c-Met-결합 FN3 도메인(0.4 nM)(P114AR7P94-A3)에 연결된 동일한 EGFR-결합 FN3 도메인을 함유하는 이중특이성 EGFR/c-Met 분자와 비교하였다. H292 및 H596 세포에서, EGFR의 인산화의 억제는 단일특이성 분자 및 이중특이성 분자(도 7의 A 및 B)에 있어서 비견되었는데, 이는 이들 세포주가 높은 EGFR 수용체 대 c-Met 수용체 비를 갖기 때문일 가능성이 높다. 이 이론을 시험하기 위하여, EGFR보다 더 많은 c-Met 수용체를 나타내는 NCI-H441 세포에서 EGFR 인산화의 억제를 평가하였다. 이중특이성 EGFR/c-Met 분자에 의한 NCI-H441 세포의 처리는, EGFR 인산화의 억제에 있어서 IC50을 단일특이성 EGFR-결합 FN3 도메인과 비교하여 30배 감소시켰다(도 7의 C).
EGFR(0.26 nM)에 대해 고 친화도를 갖고 c-Met(10.1 nM)에 대해 중간 친화도를 갖는 분자를 사용하여 c-Met 인산화 검정에서, 이중특이성 EGFR/c-Met 분자에 의한 향상된 효력에 대한 잠재성을 평가하였다. NCI-H292 세포 및 NCI-H596 세포 둘 모두에서는, 단일특이성 c-Met-결합 FN3 도메인과 비교하여 이중특이성 분자에 의해 각각 134배 및 1012배로 c-Met의 인산화의 억제를 향상시켰다(도 7의 D 및 E).
이중특이성 EGFR/c-Met 분자에 의한 EGFR 및 c-Met 인산화의 억제에 있어서의 향상된 효력이 신호전달 및 증식의 향상된 억제로 이어졌음이 검증되었다. 이들 실험을 위하여, EGFR-결합 FN3 도메인과 c-Met-결합 FN3 도메인의 혼합물을 이중특이성 EGFR/c-Met 분자와 비교하였다. 표 17 및 표 18에 기재된 바와 같이, 단일특이성 결합물질들의 혼합물과 비교하여 이중특이성 EGFR/c-Met 분자로 세포를 처리했을 때, ERK 인산화에 대한 IC50 값(표 17) 및 H292 세포의 증식(표 18)이 감소되었다. 이중특이성 EGFR/c-Met 분자의 경우의 ERK 인산화의 억제에 대한 IC50은 2개의 단일특이성 EGFR과 c-Met FN3 도메인의 혼합물에 비하여 143배 더 낮았는데, 이는 이 검정에서 이러한 분자의 효력에 대한 결합력의 효과를 나타낸다. 표 17에서, 단일특이성 EGFR 및 c-Met 결합 FN3 도메인은 활성을 완전히 억제하지 못하며, 이에 따라, 나타낸 IC50 값은 하한치인 것으로 여겨져야 한다. EGFR 결합 FN3 도메인과 c-Met 결합 FN3 도메인의 상이한 조합들을, 혼합물로서 또는 이중특이성 포맷에 연결된 것으로서 사용하여, 증식 검정을 완료하였다. 이중특이성 EGFR/c-Met 분자의 경우의 증식의 억제에 대한 IC50은 단일특이성 부모 EGFR 또는 c-Met 결합 FN3 도메인들의 혼합물에 비하여 34 내지 236배 더 낮았다. 이는 이들 수용체의 수준에서 관찰된 결합력 효과(도 6 및 도 7)가 세포 신호전달(표 17) 및 세포 증식(표 18) 억제의 개선으로 이어졌음을 확인시켜 주었다.
[표 17]
[표 18]
생체내 종양 이종이식편:PK/PD
생체내에서의 단일특이성 및 이중특이성 FN3 도메인 분자의 효능을 결정하기 위하여, 인간 HGF를 분비하도록 종양 세포를 조작하였다(뮤린 HGF는 인간 HGF에 결합하지 않는다). 렌티바이러스성 감염(진코포에이아(Genecopoeia)로부터의 인간 HGF(기탁 번호 X16322)를 발현하는 렌티바이러스 DNA 벡터 및 렌티바이러스 패키징 키트)을 이용하여, 인간 HGF를 NCI-H292 세포에서 안정하게 발현시켰다. 감염 후에, HGF-발현 세포를 4 ㎍/mL의 퓨로마이신(puromycin)(인비트로젠)을 사용하여 선택하였다. 메조 스케일 디스커버리로부터의 검정 플레이트를 사용하여 풀링된 세포의 컨디셔닝된 배지 중에서 인간 HGF 단백질을 검출하였다.
SCID 베이지 마우스에 인간 HGF를 발현하는 NCI-H292 세포(200 μL 부피의 컬트렉스(Cultrex)(트레비젠(Trevigen)) 중 2.0×106개 세포)를 각각의 동물의 등 옆부분에 피하 접종하였다. 종양 부피가 150 내지 250 ㎣의 범위가 될 때까지 매주 2회 종양 측정을 실시하였다. 이어서, (반감기를 증가시키기 위해 알부민 결합 도메인에 연결된) 이중특이성 EGFR/c-Met 분자 또는 PBS 비히클의 단회 IP 용량을 마우스에 제공하였다. 투여 후 6시간째 또는 72시간째에, 종양을 적출하고 액체 질소 중에서 즉시 냉동시켰다. 혈액 샘플을 심장 천자를 통해 프로테아제 억제제를 함유하는 3.8% 시트레이트 내로 수집하였다. 수집 직후에, 혈액 샘플을 원심분리하였으며, 생성된 혈장을 샘플관에 옮기고 -80℃에서 저장하였다. 종양의 중량을 측정하고, 작은 조각으로 절단하고, HALT 프로테아제/포스파타제 억제제(피어스(Pierce)), 50 mM 불화나트륨(시그마), 2 mM 활성화 오르토바나듐산나트륨(시그마), 및 1 mM PMSF(메조 스케일 디스커버리)와 함께 RIPA 완충액을 함유하는 용해 매트릭스 A(Lysing Matrix A) 관(LMA) 내에서 용해시켰다. 용해물을 LMA 매트릭스로부터 회수하고 원심분리하여 불용성 단백질을 제거하였다. BCA 단백질 검정을 사용하여 가용성 종양 단백질을 정량화하고, 종양 용해 완충액 중에 등가의 단백질 수준으로 희석시켰다. 메조 스케일 디스커버리로부터의 검정 플레이트를 (제조자의 프로토콜에 따라 그리고 전술된 바와 같이) 사용하여, 인산화된 c-Met, EGFR 및 ERK를 측정하였다.
도 6은 시험의 결과를 나타낸다. 각각의 이중특이성 EGFR/c-Met 분자는 6시간 및 72시간 둘 모두에서 인산화된 c-Met, EGFR, 및 ERK의 수준을 유의하게 감소시켰다. 도 6에 제시된 데이터는 c-Met 및 EGFR 둘 모두의 동시 억제의 중요성, 및 각각의 수용체에 대한 이중특이성 EGFR/c-Met 분자의 친화도가 어떻게 하류 ERK의 억제에서 역할을 하는지를 나타낸다. 고 친화도 EGFR-결합 FN3 도메인(P54AR4-83v2; 도면에서 "8"로 나타나 있음, KD=0.26 nM)을 함유하는 분자는, 6시간 및 72시간 둘 모두에서, 중간 친화도 EGFR-결합 FN3 도메인(P53A1R5-17v2; 도면에서 "17"로 나타나 있음, KD =1.9 nM)을 함유하는 분자와 비교하여 더 큰 정도로 EGFR의 인산화를 억제하였다. 시험된 4개의 이중특이성 분자 모두는 친화도에 관계 없이 6시간 시점에서 ERK의 인산화를 완전히 억제하였다. 72시간 시점에서, 고 친화도 c-Met-결합 도메인(P114AR7P94-A3; 도면에서 "A3"으로 나타나 있음, KD =0.4 nM)을 함유하는 분자는 중간 친화도 c-Met-결합 FN3 도메인(P114AR5P74-A5; 도면에서 "A5"로 나타나 있음; KD =10.1 nM; 도 6)과 비교하여 ERK의 인산화를 유의하게 억제하였다.
각각의 이중특이성 EGFR/c-Met 분자의 농도를, 투여 후 6시간째 및 72시간째에, 혈액에서 그리고 종양에서 측정하였다(도 9). 흥미롭게도, 중간 친화도 EGFR-결합 도메인(P53A1R5-17v2; KD =1.9 nM)을 갖지만 고 친화도 c-Met-결합 FN3 도메인(P114AR7P94-A3; KD =0.4 nM)을 갖는 이중특이성 분자는 다른 분자들에 비하여 6시간째에 유의하게 더 많은 종양 축적률을 가졌지만, 이 차이는 72시간쯤에는 감소되었다. 종양 밖의 세포가 EGFR 및 c-Met 둘 모두에서 더 낮은 표면 발현 수준을 가지며, 이에 따라 중간 친화도 EGFR 분자는, 더 높은 친화도의 EGFR-결합 FN3 도메인과 엄격하게 비교할 때, 정상 조직에 결합하지 않는 것으로 가설화될 수 있다. 그러므로, 종양에서 결합에 이용가능한 중간 친화도 EGFR-결합 FN3 도메인에는 더 많은 자유가 있다. 따라서, 각각의 수용체에 적절한 친화도를 확인하는 것은 감소된 전신 독성 및 증가된 종양 축적률을 갖는 치료제의 확인을 가능하게 할 수 있다.
이중특이성 EGFR/c-Met 분자에 의한 종양 효능 연구
SCID 베이지 마우스에 인간 HGF를 발현하는 NCI-H292 세포(200 μL의 컬트렉스(트레비젠) 중 2.0×106개 세포)를 각각의 동물의 등 옆부분에 피하 접종하였다. 이식 후 1주째에, 마우스를 등가의 종양 부피(평균 종양 부피=77.9+/-1.7 ㎣)를 갖는 집단으로 층별화하였다. 마우스에 이중특이성 분자를 주당 3회 투여하였으며, 종양 부피를 매주 2회 기록하였다. c-Met 및 EGFR에 대해 가변 친화도를 갖는 4개의 상이한 이중특이성 분자에 의한 종양 성장 억제(TGI)를 관찰하였다. 도 10은 종양 성장에서의 지연으로서 c-Met 및 EGFR 둘 모두의 억제 효과가, c-Met-결합 FN3 도메인이 중간 친화도였을 때, 중간 친화도 EGFR-결합 FN3 도메인과 비교하여 고 친화도 EGFR-결합 FN3 도메인을 함유하는 분자로 처리된 마우스에서 관찰되었음을 보여준다(채워진 삼각형 대비 빈 삼각형, P53A1R5-17 - P114AR5P74-A5 대비 P54AR4-83v2 - P114AR5P74-A5). 게다가, 이 데이터는 고 친화도 c-Met-결합 FN3 도메인을 갖는 것의 중요성을 보여주는데, 그 이유는 고 또는 중간 친화도 EGFR-결합 FN3 도메인을 함유하지만 고 친화도 c-Met-결합 FN3 도메인을 함유하는 이중특이성 분자가 최대 효능을 나타내었기 때문이다(회색 점선 및 흑색 점선, P54AR4-83v2 - P114AR7P94-A3 및 P53A1R5-17v2 - P114AR7P94-A3).
이중특이성 분자 및 EGFR 및 c-Met의 다른 억제제의 효능
이중특이성 EGFR/c-Met 분자(ECB38) 및 소분자 억제제인 크리조티닙(c-Met 억제제) 및 에를로티닙(EGFR 억제제), 세툭시맙(항-EGFR 항체) - 이들 각각은 단제(single agent)로서임 -, 및 크리조티닙과 에를로티닙의 병용의 생체내 치료적 효능을 SCID/베이지 마우스에서의 피하 H292-HGF 인간 폐암 이종이식편 모델의 처리에서 평가하였다(도 11).
H292-HGF 세포를 공기 중 5% CO2의 분위기 내에서 37℃에서 소태아 혈청(10% v/v) 및 L-글루타민(2 mM)으로 보충된 RPMI1640 배지 중에 시험관내에서 유지하였다. 트립신-EDTA 처리에 의해 세포를 매주 2회 일상적으로 계대배양하였다. 지수 성장기에서 성장하는 세포를 수거하고 종양 접종을 위해 카운팅하였다.
종양 발생을 위하여, 컬트렉스를 함유하는 PBS(1:1) 0.1 ml 중 H292-HGF 종양 세포(2 × 106)를 각각의 마우스의 우측 옆구리 부분에 피하 접종하였다. 평균 종양 크기가 139 ㎣에 도달했을 때 이 처리를 시작하였다. 각각의 연구 집단에서의 시험 대상품(test article) 투여 및 동물 마리수는 하기 실험 설계 표(표 26)에 나타나 있다. 종양 세포 접종 일자는 일수 0으로서 표기하였다.
[표 26]
치료 개시 전에, 모든 동물의 체중을 측정하고 종양 부피를 측정하였다. 종양 부피는 임의의 주어진 치료의 유효성에 영향을 줄 수 있기 때문에, 그들의 종양 부피에 기초한 무작위화 블록 설계(randomized block design)를 사용하여 마우스를 집단에 배정하였다. 이는 모든 집단이 기준선에서 비견되도록 보장한다. 무작위화 블록 설계를 사용하여 실험 동물들을 집단에 배정하였다. 첫 번째로, 실험 동물들을 그들의 초기 종양 부피에 따라 균질한 블록들로 나누었다. 두 번째로, 각각의 블록 내에서, 치료에 대한 실험 동물들의 무작위화를 수행하였다. 무작위화 블록 설계를 사용하여 실험 동물들을 배정함으로써, 각각의 동물이 주어진 치료에 배정되는 데 동일한 확률을 가지며, 이에 따라 계통 오류(systematic error)를 감소시키도록 보장하였다.
일상적인 모니터링시에, 움직임, 먹이 및 물 소비의 시각적 평가, 체중 증가/손실(체중은 매주 2회 측정하였음), 눈/모발의 윤기 상실(matting) 및 임의의 다른 비정상 효과와 같은, 정상 거동에 대한 종양 성장 및 치료의 어떠한 효과에 대해서도 동물들을 검사하였다.
주요 종점은 종양 성장이 지연될 수 있는지 또는 종양 보유 마우스가 치유될 수 있는지 중 어느 것이었다. 캘리퍼를 사용하여 종양 크기를 2개의 치수에서 매주 2회 측정하였으며, 부피는 하기 식을 사용하여 ㎣로 표현하였다: V = 0.5 a × b 2(여기서, a 및 b는 각각 종양의 긴 직경 및 짧은 직경임). 이어서, 종양 크기를 T-C 및 T/C 값 둘 모두의 계산에 사용하였다. 처리군의 평균 종양 크기가 1000 ㎣에 도달하는 데 필요한 시간(단위: 일수)으로서 T를 사용하여 T-C를 계산하였으며, C는 대조군의 평균 종양 크기가 동일한 크기에 도달하는 데 걸린 시간(단위: 일수)이었다. T/C 값(단위: %)은 항종양 효능의 지표였으며; T 및 C는 주어진 날에 대한 각각 처리군 및 대조군의 평균 부피였다. 완전 종양 퇴행(complete tumor regression, CR)은 촉진(palpation)의 한계(62.5 ㎣) 미만으로 감소된 종양으로서 규정된다. 부분 종양 퇴행(partial tumor regression, PR)은 초기 종양 부피로부터 감소된 종양으로서 규정된다. 3회 이상의 연속 종양 측정에서의 CR 또는 PR의 최소한의 지속시간이, CP 또는 PR을 지속적이라고 간주하는 데 필요하다.
체중 손실이 20%를 초과하거나 또는 집단의 평균 종양 크기가 2000 ㎣를 초과하는 동물들을 안락사시켰다. 이 연구는 최종 투여 후 2주의 관찰 후에 종료하였다.
평균 및 평균의 표준 오차(SEM)를 포함하는 요약 통계치가 각각의 시점에서 각각의 집단의 종양 부피에 대해 제공된다(하기 표 19에 나타냄). 집단들 사이의 종양 부피 차이의 통계학적 분석을 일원분산분석(one-way ANOVA)을 사용하여 평가한 후, 게임즈-호웰(Games-Howell)(등분산성이 가정되지 않음)을 사용하여 개별 비교들을 행하였다. 모든 데이터는 SPSS 18.0을 사용하여 분석하였다. p< 0.05를 통계학적으로 유의한 것으로 간주하였다.
[표 19]
비히클 처리군(집단 1)의 평균 종양 크기는 종양 접종 후 일수 23에서 1,758 ㎣에 도달하였다. 25 mg/㎏ 용량 수준의 이중특이성 EGFR/c-Met 분자(집단 2)에 의한 처리는 모든 마우스에서 완전 종양 퇴행(CR)으로 이어졌는데, 이때 이는 3회 초과의 연속 종양 측정에서 지속적이었다(일수 23의 비히클군과 비교하여 평균 TV=23 ㎣, T/C 값=1%, p=0.004).
50 mg/㎏ 용량 수준의 단제로서의 크리조티닙(집단 3)에 의한 치료는 항종양 활성을 나타내지 않았으며; 평균 종양 크기는 일수 23에서 2,102 ㎣였다(비히클군과 비교하여 T/C 값=120%, p=0.944).
50 mg/㎏ 용량 수준의 단제로서의 에를로티닙(집단 4)에 의한 치료는 미소한 항종양 활성을 나타내었지만, 비히클군과 비교하여 유의한 차이는 발견되지 않았으며; 평균 종양 크기는 일수 23에서 1,122 ㎣였는데(비히클군과 비교하여 T/C 값=64%, p=0.429), 이때 비히클군과 비교하여 1,000 ㎣의 종양 크기에서 4일의 종양 성장 지연이 있었다.
크리조티닙(50 mg/㎏, 집단 5)과 에를로티닙(50 mg/㎏, 집단 5)의 병용은 유의한 항종양 활성을 나타내었으며; 평균 종양 크기는 일수 23에서 265 ㎣였는데(T/C=15%; p=0.008), 이때 비히클군과 비교하여 1,000 ㎣의 종양 크기에서 17일의 종양 성장 지연이 있었다.
단제로서의 1 mg/마우스 용량 수준의 세툭시맙(집단 6)은 유의한 항종양 활성을 나타내었으며; 평균 종양 크기는 일수 23에서 485 ㎣였는데(T/C=28%; p=0.018), 이때 비히클군과 비교하여 1,000 ㎣의 종양 크기에서 17일의 종양 성장 지연이 있었다. 도 11은 다양한 요법들의 항종양 활성을 나타낸다.
[표 20]
중간 내지 심각한 체중 손실이 비히클군에서 관찰되었는데, 이는 증가하는 종양 부하(tumor burden)에 기인할 수 있었을 것이며; 3 마리는 죽었으며, 1 마리의 마우스는 일수 23일까지 BWL>20%이었을 때 안락사시켰다. 이중특이성 EGFR/c-Met 분자의 약간의 독성이 집단 2에서 관찰되었는데; 3 마리의 마우스를 처리 기간 동안 BWL>20%일 때 안락사시켰으며; 2주의 관찰 기간 동안 처리를 중단했을 때 체중을 점차로 회복하였다. 비히클군과 비교하여 크리조티닙 또는 에를로티닙의 단제요법군에서 더 심각한 체중 손실이 관찰되었는데, 이는 치료 관련 독성을 시사한다. 크리조티닙과 에를로티닙의 병용은 일반적으로 투여 단계 동안 내약성을 나타내었지만, 연구의 종료시에 심각한 체중 손실이 관찰되었는데, 이는 비처리 기간 동안의 빠른 종양 성장의 재개에 기인할 수 있었을 것이다. 세툭시맙의 단제요법은 이 연구에서 우수한 내약성을 나타내었으며; 체중 손실은 종양 성장의 재개로 인해 단지 연구의 종료시에 관찰되었다.
요약하면, 25 mg/㎏(3회/주 × 3주)의 이중특이성 EGFR/c-Met 분자는 SCID/베이지 마우스에서 H292-HGF 인간 폐암 이종이식편 모델에서 완전한 반응을 생성하였다. 이 처리는 10 마리의 마우스 중 7 마리에서 내약성을 나타내었는데, 결과적으로는 10 마리의 마우스 중 3 마리에서 심각한 체중 손실이 있었다. 도 11 및 표 20은 처리 후 시점들 동안의 종양 크기에 대한 다양한 요법들의 영향을 나타낸다.
실시예 9: 이중특이성 EGFR/c-Met 분자의 반감기 연장
신장 여과를 감소시키고 이에 따라 단백질의 혈청 반감기를 연장하기 위해 다수의 방법이 기술되어 왔는데, 이러한 방법은 폴리에틸렌 글리콜(PEG) 또는 다른 중합체에 의한 변형, 알부민에 대한 결합, 알부민 또는 다른 혈청 단백질에 결합하는 단백질 도메인에 대한 융합, 알부민에 대한 유전자 융합, IgG Fc 도메인에 대한 융합, 및 긴 비구조화 아미노산 서열에 대한 융합을 포함한다.
분자의 C-말단에 유리 시스테인을 도입시킴으로써 수력학적 반경을 증가시키기 위하여 이중특이성 EGFR/c-Met 분자를 PEG로 변형시켰다. 가장 일반적으로는, 시스테인 잔기의 유리 티올 기를 사용하여, 표준 방법을 사용하여 말레이미드 또는 요오도아세트아미드 기로 작용화된 PEG 분자를 부착한다. 1000, 2000, 5000, 10,000, 20,000, 또는 40,000 kDa의 선형 PEG를 포함하는 다양한 형태의 PEG를 사용하여 단백질을 변형시킬 수 있다. 이들 분자량의 분지형 PEG 분자가 또한 변형에 사용될 수 있다. 일부 경우에, PEG 기는 또한 이중특이성 EGFR/c-Met 분자 내의 1차 아민을 통해 부착될 수 있다.
페길화(PEGylation)에 더하여, 천연 발생 3-나선 번들 혈청 알부민 결합 도메인(ABD) 또는 공통 알부민 결합 도메인(ABDCon)과의 융합 분자로서 이들 단백질을 생성함으로써 이중특이성 EGFR/c-Met 분자의 반감기를 연장시켰다. 이들 단백질 도메인을 표 16에 기재된 링커들 중 임의의 것을 통해 c-Met-결합 FN3 도메인의 C-말단에 연결시켰다. ABD 또는 ABDCon 도메인은 또한 1차 서열 내의 EGFR-결합 FN3 도메인과 c-Met 결합 FN3 도메인 사이에 배치될 수 있다.
실시예 10: 선택된 이중특이성 EGFR/c-Met 분자의 특성화
선택된 EGFR/c-Met 분자들을, EGFR 및 c-Met 둘 모두에 대한 그들의 친화도, EGFR 및 c-Met 자가인산화를 억제하는 그들의 능력, 및 HGF 세포의 증식에 대한 그들의 영향에 대해 특성화하였다. 재조합 EGFR 및/또는 c-Met 세포외 도메인에 대한 이중특이성 EGFR/c-Met 분자의 결합 친화도는 실시예 3에 기재된 프로토콜에 따라 프로테온 기기(바이오라드)를 사용하여 추가로 표면 플라즈몬 공명 방법에 의한 것이었다. 특성화의 결과가 표 21에 나타나 있다.
[표 21]
실시예 11: 시스테인 조작된 이중특이성 항-EGFR/c-Met 분자의 생성 및 특성화
이중특이성 EGFR/c-Met 분자의 생성
P54AR4-83v2 돌연변이체(실시예 5)의 시스테인 스캐닝으로부터 생성된 데이터에 기초하여, 시스테인 돌연변이체를 또한 ECB147(서열 번호 218 및 256)로 표기된 이중특이성 항-EGFR/c-Met 분자 내에 설계하였는데, 이 분자는 P54AR4-83v2(서열 번호 27), cMet 결합제 P114AR7P95-C5v2(서열 번호 114), 및 반감기 연장을 위한 알부민 결합 도메인으로 이루어진다. 이들 3개의 도메인은 (Ala-Pro)5 링커(서열 번호 81)에 의해 연결된다. 1개, 2개, 또는 4개의 시스테인을 갖는 변이체들을 FN3 도메인들(서열 번호 219-225 및 257-263) 중 하나의 C-말단에서, 링커 영역 내에서, 또는 Lys-62 위치에서 치환에 의해 설계하였다. 다른 이중특이성 변이체, ECB82cys(서열 번호 226 및 264)는 P54AR4-83v2(서열 번호 27), P114AR7P94-A3v22(서열 번호 111), 및 알부민-결합 도메인의 변이체로 이루어지며, 3개의 모든 도메인은 AP 링커, 및 단일 C-말단 시스테인에 의해 연결되어 있다. 비-표적화된 텐콘 스캐폴드(서열 번호 265)의 추가의 시스테인 변이체를 또한 대조 접합체의 작제에 사용하였다. 모든 변이체를 앞서의 실시예에 기재된 바와 같이 작제하고, 발현하고, 정제하였다. SDS-PAGE 분석에 의해 순도를 평가하였다. 수퍼덱스 75 5/150 컬럼(지이 헬쓰케어)을 사용하는 분석용 크기 배제 크로마토그래피는 FN3 도메인 조제물에 응집체가 없고 단량체성 단백질과 일치되게 일시에 용출됨을 보여준다. 질량 분석에 의해 질량이 이론상의 질량과 일치하는 것으로 판단되었다(표 22).
[표 22]
화학적 접합
정제된 이중특이성 시스테인 변이체를 말레이미드-함유 분자에 화학적으로 접합시키기 위하여, 단백질을 먼저 TCEP로 환원시켜 유리 티올을 생성하였다. 1 내지 2 mg의 각각의 이중특이성 시스테인 변이체를 중성 pH의 과량의 TCEP(시그마 카탈로그 번호 646547)와 혼합하고 실온에서 30 내지 60분 동안 인큐베이션하였다. 3 부피의 포화 황산암모늄 용액(4.02 M)을 첨가하여 시스테인 변이체를 침전시킴으로써 TCEP를 제거하였다. 4℃에서 20분 동안 16000 내지 20000 × g로 원심분리하고 상청액을 제거한 후에, 단백질 펠릿을 PBS 또는 인산나트륨 완충액 중에 용해시키고 5 내지 10배 과량의 말레이미드-함유 분자와 즉시 혼합하였다. 반응물을 실온에서 30 내지 60분 동안 인큐베이션하고, 이어서 과량의 유리 티올, 예컨대 시스테인 또는 β-메르캅토에탄올로 켄칭하여(quench), 과량의 말레이미드를 포착하였다. 결합되지 않은 말레이미드를, 토소(Tosoh) G3000SWxl 컬럼(번호 P4619-14N; 7.8 mm × 30 cm; 5 μm)을 갖는 분취용 SEC에 의해 제바(Zeba) 탈염 컬럼(써모(Thermo) 카탈로그 번호 89890)으로 제거하거나, 또는 본질적으로 전술된 바와 같이 시스테인 변이체를 Ni-NTA 수지에 결합하고, 세척하고, 용출시킴으로써 제거하였다. SDS-PAGE 및 질량 분석에 의해 접합체를 특성화하였다. 이러한 일반적 방법을 사용하여 이중특이성 시스테인 변이체를 플루오레세인 말레이미드(써모 카탈로그 번호 62245), PEG24-말레이미드(퀀타 바이오디자인(Quanta Biodesign) 카탈로그 번호 10319), 및 말레이미드-세포독소 분자에 다양한 링커를 사용하여 접합시켰다(도 2에서의 구조 참조).
EGF-자극된 EGFR 인산화의 억제
메조 스케일 디스커버리(미국 매릴랜드주 게이더스버그 소재)로부터의 EGFR 포스포(Tyr1173) 키트를 사용하여 그리고 실시예 3에 기재된 바와 같이, 정제된 이중특이성 PEG24-말레이미드 접합체들을 인간 종양 세포주 NCI-H292(ATCC, 카탈로그 번호 CRL-1848)에서의 EGFR의 EGF-자극된 인산화를 억제하는 그들의 능력에 대해 시험하였다. 접합체들을 비변형 ECB38(서열 번호 109)과 대비하였는데, 이때 비변형 ECB38은 ECB147과 2개의 아미노산만큼 상이하다. 접합체들 및 ECB38은 표 23에 나타낸 바와 같이 유사한 IC50 값으로 EGFR을 억제하였는데, 이는 설계된 부위에서의 변형이 표적 결합에 유의하게 영향을 주지 않음을 입증한다.
[표 23]
HGF-자극된 c-Met 인산화의 억제
메조 스케일 디스커버리(미국 매릴랜드주 게이더스버그 소재)로부터의 c-Met 포스포(Tyr1349) 키트를 사용하여, 그리고 실시예 7에 기재된 바와 같이, 정제된 이중특이성 PEG24-말레이미드 접합체들을 또한 NCI-H292 세포에서의 c-Met의 HGF-자극된 인산화를 억제하는 그들의 능력에 대해 시험하였다. 접합체들 및 ECB38은 표 24에 나타낸 바와 같이 유사한 IC50 값으로 cMet를 억제하였는데, 이는 이들 부위에서의 변형이 표적 결합을 유의하게 변경시키지 않음을 입증한다.
[표 24]
세포독성 검정
오리스타틴 패밀리로부터의 세포독성 튜불린 억제제에 연결된 ECB147 시스테인 변이체, 83v2-cys, 또는 텐콘-cys로 이루어진 접합체들(도 2)을 암 세포에서 표적-의존성 세포독성에 대해 시험하였다. 이 억제제를 절단 불가능 PEG4 링커 또는 효소-절단가능 발린-시트룰린 또는 발린-라이신 링커를 통해 시스테인-함유 단백질에 연결하였다. 실시예 4에 기재된 절차를 사용하여 단백질-세포독소 접합체에 노출시킨 후에 EGFR-과발현 인간 종양 세포주 H1573 및 A431뿐만 아니라 EGFR-음성 종양 세포주 MDA-MB-435의 생존력을 측정함으로써 세포 사멸을 평가하였다. 표 25에는 66, 72, 또는 90시간의 시점에서의 셀타이터 글로 또는 인큐사이트(IncuCyte) 객체 카운트 데이터의 분석으로부터 얻어진 IC50 값이 기록되어 있다. 단백질-약물 접합체는 표적 항원 EGFR을 발현하는 세포에 대해 강력한 세포-사멸을 보여주었다. 다중-약물 접합체가 또한 시험된 세포주들 중 다수에서 증가된 세포독성을 입증하였다.
[표 25]
서열 목록
서열 번호 73, PRT, 호모 사피엔스(Homo Sapiens), EGFR
1 mrpsgtagaa llallaalcp asraleekkv cqgtsnkltq lgtfedhfls lqrmfnncev
61 vlgnleityv qrnydlsflk tiqevagyvl ialntverip lenlqiirgn myyensyala
121 vlsnydankt glkelpmrnl qeilhgavrf snnpalcnve siqwrdivss dflsnmsmdf
181 qnhlgscqkc dpscpngscw gageencqkl tkiicaqqcs grcrgkspsd cchnqcaagc
241 tgpresdclv crkfrdeatc kdtcpplmly npttyqmdvn pegkysfgat cvkkcprnyv
301 vtdhgscvra cgadsyemee dgvrkckkce gpcrkvcngi gigefkdsls inatnikhfk
361 nctsisgdlh ilpvafrgds fthtppldpq eldilktvke itgflliqaw penrtdlhaf
421 enleiirgrt kqhgqfslav vslnitslgl rslkeisdgd viisgnknlc yantinwkkl
481 fgtsgqktki isnrgensck atgqvchalc spegcwgpep rdcvscrnvs rgrecvdkcn
541 llegeprefv enseciqchp eclpqamnit ctgrgpdnci qcahyidgph cvktcpagvm
601 genntlvwky adaghvchlc hpnctygctg pglegcptng pkipsiatgm vgalllllvv
661 algiglfmrr rhivrkrtlr rllqerelve pltpsgeapn qallrilket efkkikvlgs
721 gafgtvykgl wipegekvki pvaikelrea tspkankeil deayvmasvd nphvcrllgi
781 cltstvqlit qlmpfgclld yvrehkdnig sqyllnwcvq iakgmnyled rrlvhrdlaa
841 rnvlvktpqh vkitdfglak llgaeekeyh aeggkvpikw malesilhri ythqsdvwsy
901 gvtvwelmtf gskpydgipa seissilekg erlpqppict idvymimvkc wmidadsrpk
961 freliiefsk mardpqrylv iqgdermhlp sptdsnfyra lmdeedmddv vdadeylipq
1021 qgffsspsts rtpllsslsa tsnnstvaci drnglqscpi kedsflqrys sdptgalted
1081 siddtflpvp eyinqsvpkr pagsvqnpvy hnqplnpaps rdphyqdphs tavgnpeyln
1141 tvqptcvnst fdspahwaqk gshqisldnp dyqqdffpke akpngifkgs taenaeylrv
1201 apqssefiga
서열 번호 75, PRT, 호모 사피엔스, 테나신-C
1 mgamtqllag vflaflalat eggvlkkvir hkrqsgvnat lpeenqpvvf nhvyniklpv
61 gsqcsvdles asgekdlapp sepsesfqeh tvdgenqivf thriniprra cgcaaapdvk
121 ellsrleele nlvsslreqc tagagcclqp atgrldtrpf csgrgnfste gcgcvcepgw
181 kgpncsepec pgnchlrgrc idgqcicddg ftgedcsqla cpsdcndqgk cvngvcicfe
241 gyagadcsre icpvpcseeh gtcvdglcvc hdgfagddcn kplclnncyn rgrcvenecv
301 cdegftgedc selicpndcf drgrcingtc yceegftged cgkptcphac htqgrceegq
361 cvcdegfagv dcsekrcpad chnrgrcvdg rcecddgftg adcgelkcpn gcsghgrcvn
421 gqcvcdegyt gedcsqlrcp ndchsrgrcv egkcvceqgf kgydcsdmsc pndchqhgrc
481 vngmcvcddg ytgedcrdrq cprdcsnrgl cvdgqcvced gftgpdcael scpndchgqg
541 rcvngqcvch egfmgkdcke qrcpsdchgq grcvdgqcic hegftgldcg qhscpsdcnn
601 lgqcvsgrci cnegysgedc sevsppkdlv vtevteetvn lawdnemrvt eylvvytpth
661 egglemqfrv pgdqtstiiq elepgveyfi rvfailenkk sipvsarvat ylpapeglkf
721 ksiketsvev ewdpldiafe tweiifrnmn kedegeitks lrrpetsyrq tglapgqeye
781 islhivknnt rgpglkrvtt trldapsqie vkdvtdttal itwfkplaei dgieltygik
841 dvpgdrttid ltedenqysi gnlkpdteye vslisrrgdm ssnpaketft tgldaprnlr
901 rvsqtdnsit lewrngkaai dsyrikyapi sggdhaevdv pksqqattkt tltglrpgte
961 ygigvsavke dkesnpatin aateldtpkd lqvsetaets ltllwktpla kfdryrlnys
1021 lptgqwvgvq lprnttsyvl rglepgqeyn vlltaekgrh kskparvkas teqapelenl
1081 tvtevgwdgl rlnwtaadqa yehfiiqvqe ankveaarnl tvpgslravd ipglkaatpy
1141 tvsiygviqg yrtpvlsaea stgetpnlge vvvaevgwda lklnwtapeg ayeyffiqvq
1201 eadtveaaqn ltvpgglrst dlpglkaath ytitirgvtq dfsttplsve vlteevpdmg
1261 nltvtevswd alrlnwttpd gtydqftiqv qeadqveeah nltvpgslrs meipglragt
1321 pytvtlhgev rghstrplav evvtedlpql gdlavsevgw dglrlnwtaa dnayehfviq
1381 vqevnkveaa qnltlpgslr avdipgleaa tpyrvsiygv irgyrtpvls aeastakepe
1441 ignlnvsdit pesfnlswma tdgifetfti eiidsnrlle tveynisgae rtahisglpp
1501 stdfivylsg lapsirtkti satattealp llenltisdi npygftvswm asenafdsfl
1561 vtvvdsgkll dpqeftlsgt qrklelrgli tgigyevmvs gftqghqtkp lraeivteae
1621 pevdnllvsd atpdgfrlsw tadegvfdnf vlkirdtkkq sepleitlla pertrditgl
1681 reateyeiel ygiskgrrsq tvsaiattam gspkevifsd itensatvsw raptaqvesf
1741 rityvpitgg tpsmvtvdgt ktqtrlvkli pgveylvsii amkgfeesep vsgsfttald
1801 gpsglvtani tdsealarwq paiatvdsyv isytgekvpe itrtvsgntv eyaltdlepa
1861 teytlrifae kgpqksstit akfttdldsp rdltatevqs etalltwrpp rasvtgyllv
1921 yesvdgtvke vivgpdttsy sladlspsth ytakiqalng plrsnmiqti fttigllypf
1981 pkdcsqamln gdttsglyti ylngdkaeal evfcdmtsdg ggwivflrrk ngrenfyqnw
2041 kayaagfgdr reefwlgldn lnkitaqgqy elrvdlrdhg etafavydkf svgdaktryk
2101 lkvegysgta gdsmayhngr sfstfdkdtd saitncalsy kgafwyrnch rvnlmgrygd
2161 nnhsqgvnwf hwkghehsiq faemklrpsn frnlegrrkr a
>서열 번호 101
PRT
호모 사피엔스
cMet
1 mkapavlapg ilvllftlvq rsngeckeal aksemnvnmk yqlpnftaet piqnvilheh
61 hiflgatnyi yvlneedlqk vaeyktgpvl ehpdcfpcqd csskanlsgg vwkdninmal
121 vvdtyyddql iscgsvnrgt cqrhvfphnh tadiqsevhc ifspqieeps qcpdcvvsal
181 gakvlssvkd rfinffvgnt inssyfpdhp lhsisvrrlk etkdgfmflt dqsyidvlpe
241 frdsypikyv hafesnnfiy fltvqretld aqtfhtriir fcsinsglhs ymemplecil
301 tekrkkrstk kevfnilqaa yvskpgaqla rqigaslndd ilfgvfaqsk pdsaepmdrs
361 amcafpikyv ndffnkivnk nnvrclqhfy gpnhehcfnr tllrnssgce arrdeyrtef
421 ttalqrvdlf mgqfsevllt sistfikgdl tianlgtseg rfmqvvvsrs gpstphvnfl
481 ldshpvspev ivehtlnqng ytlvitgkki tkiplnglgc rhfqscsqcl sappfvqcgw
541 chdkcvrsee clsgtwtqqi clpaiykvfp nsapleggtr lticgwdfgf rrnnkfdlkk
601 trvllgnesc tltlsestmn tlkctvgpam nkhfnmsiii snghgttqys tfsyvdpvit
661 sispkygpma ggtlltltgn ylnsgnsrhi siggktctlk svsnsilecy tpaqtistef
721 avklkidlan retsifsyre dpivyeihpt ksfistwwke plnivsflfc fasggstitg
781 vgknlnsvsv prmvinvhea grnftvacqh rsnseiicct tpslqqlnlq lplktkaffm
841 ldgilskyfd liyvhnpvfk pfekpvmism gnenvleikg ndidpeavkg evlkvgnksc
901 enihlhseav lctvpndllk lnselniewk qaisstvlgk vivqpdqnft gliagvvsis
961 talllllgff lwlkkrkqik dlgselvryd arvhtphldr lvsarsvspt temvsnesvd
1021 yratfpedqf pnssqngscr qvqypltdms piltsgdsdi sspllqntvh idlsalnpel
1081 vqavqhvvig psslivhfne vigrghfgcv yhgtlldndg kkihcavksl nritdigevs
1141 qfltegiimk dfshpnvlsl lgiclrsegs plvvlpymkh gdlrnfirne thnptvkdli
1201 gfglqvakgm kylaskkfvh rdlaarncml dekftvkvad fglardmydk eyysvhnktg
1261 aklpvkwmal eslqtqkftt ksdvwsfgvl lwelmtrgap pypdvntfdi tvyllqgrrl
1321 lqpeycpdpl yevmlkcwhp kaemrpsfse lvsrisaifs tfigehyvhv natyvnvkcv
1381 apypsllsse dnaddevdtr pasfwets
>서열 번호 179
PRT
인공
EGFR 결합 FN3 도메인의 FG 루프
HNVYKDTNX9RGL
(여기서, X9는 M 또는 I임)
>서열 번호 180
PRT
인공
EGFR 결합 FN3 도메인의 FG 루프
LGSYVFEHDVML(서열 번호 180),
>서열 번호 181
PRT
인공
EGFR 결합 FN3 도메인의 BC 루프
X1X2X3X4X5X6X7X8(서열 번호 181)
(여기서,
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L임)
>서열 번호 182
PRT
인공
EGFR 결합 FN3 도메인
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX9RGLPLSAEFTT(서열 번호 182)
(X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L이고;
X9는 M 또는 I임)
>서열 번호 183
PRT
인공
EGFR 결합 FN3 도메인
LPAPKNLVVSEVTEDSLRLSWX1X2X3X4X5X6X7X8DSFLIQYQESEKVGEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVLGSYVFEHDVMLPLSAEFTT(서열 번호 183)
(여기서,
X1은 A, T, G 또는 D이고;
X2는 A, D, Y 또는 W이고;
X3은 P, D 또는 N이고;
X4는 L이거나 또는 부재하고;
X5는 D, H, R, G, Y 또는 W이고;
X6은 G, D 또는 A이고;
X7은 A, F, G, H 또는 D이고;
X8은 Y, F 또는 L임)
>서열 번호 184
PRT
인공
c-Met 결합 FN3 도메인의 C 가닥 및 CD 루프 서열
DSFX10IRYX11E X12X13X14X15GX16(서열 번호 184)
(여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D임)
>서열 번호 185
PRT
인공
c-Met 결합 FN3 도메인의 F 가닥 및 FG 루프
TEYX17VX18IX19X20V KGGX21X22SX23(서열 번호 185)
(여기서,
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L임)
>서열 번호 186
PRT
인공
c-Met 결합 FN3 도메인
LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX10IRYX11EX12X13X14X15GX16AIVLTVPGSERSYDLTGLKPGTEYX17VX18IX19X20VKGGX21X22SX23PLSAEFTT(서열 번호 186)
(여기서,
X10은 W, F 또는 V이고;
X11은 D, F 또는 L이고;
X12는 V, F 또는 L이고;
X13은 V, L 또는 T이고;
X14는 V, R, G, L, T 또는 S이고;
X15는 G, S, A, T 또는 K이고;
X16은 E 또는 D이고;
X17은 Y, W, I, V, G 또는 A이고;
X18은 N, T, Q 또는 G이고;
X19는 L, M, N 또는 I이고;
X20은 G 또는 S이고;
X21은 S, L, G, Y, T, R, H 또는 K이고;
X22는 I, V 또는 L이고;
X23은 V, T, H, I, P, Y, T 또는 L임)
>서열 번호 187
PRT
인공
이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 EGFR FN3 도메인
LPAPKNLVVSX24VTX25DSX26RLSWDDPX27AFYX28SFLIQYQX29SEKVGEAIX30LTVPGSERSYDLTGLKPGTEYTVSIYX31VHNVYKDTNX32RGLPLSAX33FTT(서열 번호 187)
(여기서,
X24는 E, N 또는 R이고;
X25는 E 또는 P이고;
X26은 L 또는 A이고;
X27은 H 또는 W이고;
X28은 E 또는 D이고;
X29는 E 또는 P이고;
X30은 N 또는 V이고;
X31은 G 또는 Y이고;
X32는 M 또는 I이고;
X33은 E 또는 I임)
>서열 번호 188
이중특이성 EGFR/c-Met FN3 도메인 함유 분자의 c-Met FN3 도메인
LPAPKNLVVSX34VTX35DSX36RLSWTAPDAAFDSFWIRYFX37FX38X39X40GX41AIX42LTVPGSERSYDLTGLKPGTEYVVNIX43X44VKGGX45ISPPLSAX46FTT(서열 번호 188)
(여기서,
X34는 E, N 또는 R이고;
X35는 E 또는 P이고;
X36은 L 또는 A이고;
X37은 E 또는 P이고;
X38은 V 또는 L이고;
X39는 G 또는 S이고;
X40은 S 또는 K이고;
X41은 E 또는 D이고;
X42는 N 또는 V이고;
X43은 L 또는 M이고;
X44는 G 또는 S이고;
X45는 S 또는 K이고;
X46은 E 또는 I임)