JP2024502837A - 高病原性コロナウイルスに対する細胞傷害性t細胞免疫療法 - Google Patents
高病原性コロナウイルスに対する細胞傷害性t細胞免疫療法 Download PDFInfo
- Publication number
- JP2024502837A JP2024502837A JP2023541012A JP2023541012A JP2024502837A JP 2024502837 A JP2024502837 A JP 2024502837A JP 2023541012 A JP2023541012 A JP 2023541012A JP 2023541012 A JP2023541012 A JP 2023541012A JP 2024502837 A JP2024502837 A JP 2024502837A
- Authority
- JP
- Japan
- Prior art keywords
- coronavirus
- antigen
- cov
- sars
- protein
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images













Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/215—Coronaviridae, e.g. avian infectious bronchitis virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5258—Virus-like particles
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/57—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2
- A61K2039/572—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2 cytotoxic response
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/70—Multivalent vaccine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/14011—Baculoviridae
- C12N2710/14023—Virus like particles [VLP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/14011—Baculoviridae
- C12N2710/14034—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/20011—Papillomaviridae
- C12N2710/20023—Virus like particles [VLP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20034—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20071—Demonstrated in vivo effect
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Virology (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Chemical & Material Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Communicable Diseases (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Mycology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Oncology (AREA)
- Molecular Biology (AREA)
- Epidemiology (AREA)
- Immunology (AREA)
- Microbiology (AREA)
- Organic Chemistry (AREA)
- Pulmonology (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicinal Preparation (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract

病原性コロナウイルスおよび感冒コロナウイルスに対する細胞傷害性Tリンパ球(CD8+)応答、すなわちMHCクラスI拘束性T細胞応答、を誘導する組成物および方法であって、パピローマウイルスL1タンパク質のHIループに挿入されたシステイン残基で挟まれた連続した負電荷アミノ酸を有するポリイオンパピローマウイルスのウイルス様粒子(VLP)からなる抗原の送達プラットフォームを含む。VLPと対になる抗原は、病原性コロナウイルス由来の融合ペプチド/タンパク質であって、MHCクラスIエピトープの提示を増強するために、システイン残基およびC末端タンパク質プロセシング配列(AAYY)が前および/または後に続く連続する正電荷アミノ酸からなるN末端またはC末端のアミノ酸を有する、ヒト集団で一般的に循環している遺伝的に最も近縁のヒトコロナウイルス由来の融合ペプチド/タンパク質、を含む。
Description
本発明は、コロナウイルスに対する細胞性免疫応答を誘導するための組成物および方法に関する。
コロナウイルス(CoV)は、サイズが約26~32キロベースの一本鎖プラスセンスRNAゲノムを有するエンベロープウイルスである。SARS-CoV-2は、他の全てのコロナウイルスと同様にそのゲノムの構成と発現に共通の特徴を有しており、オープンリーディングフレーム(ORF)1a/bによってコードされる非構造タンパク質の後に、主要な構造タンパク質スパイク(S)、エンベロープ(E)、膜(M)、およびヌクレオカプシド(N)が続く(Chenら、2020年)。S糖タンパク質は、宿主受容体への結合を媒介する。S糖タンパク質は、宿主プロテアーゼによって、S1およびS2と称される2つの別々のポリペプチドに切断される。S1は、Sタンパク質の受容体結合ドメインを構成し、中和抗体の主要な標的であるが、S2は、スパイク分子のストーク(柄)を形成する。Mタンパク質は、最も豊富なウイルスタンパク質であり、コアウイルス粒子の形成に関与する。Nタンパク質は、唯一のヌクレオカプシドタンパク質であり、ウイルスゲノムと相互作用するコア粒子の部分を形成する。CoVは、系統発生に基づいて、α-CoV、β-CoV、γ-CoV、およびδ-CoVの4つの属に分類される。β-CoV属内では、4つの系統(A、B、C、およびD)が認識されている。
コロナウイルスは、動物に多種多様な疾患を引き起こす。ヒトの場合、CoV感染は主に上気道および胃腸管に影響を及ぼし、主に、風邪などの軽度の自然治癒性の疾患を引き起こす。これらのヒトコロナウイルスのうち、2つはα-コロナウイルスであるHCoV-229EおよびHCoV-NL63であり、他の2つはβ-コロナウイルスであるHCoV-OC43およびHCoV-HKU1である。HCoV-229EおよびHCoV-OC43は略50年前に単離されたが、HCoV-NL63およびHCoV-HKU1は最近になって同定されたに過ぎない。これらのウイルスはヒト集団に常在しており、毎年呼吸器感染症の15~30%を引き起こしている。血清有病率調査から、これらのウイルスへの曝露はヒトにおいてほぼ普遍的であることが示唆されている(Severance et al、2008)。さらなるヒトコロナウイルスの存在も考えられるが、不明である。最初の高病原性ヒトコロナウイルスは、β-コロナウイルスであるSARS-CoVであった。中国の広東省で2002~2003年に発生した重症急性呼吸器症候群(SARS)の原因物質として同定された。2012年に中東で、2番目の新規高病原性ヒトCoVが出現した。中東呼吸器症候群-CoV(MERS-CoV)と名付けられたこのウイルスもβ-コロナウイルスでもあり、サウジアラビアをはじめとする中東諸国における高病原性呼吸器感染症の原因物質であることが判明した。2019年後半に中国で初めて出現した、3番目で最新の高病原性ヒトコロナウイルスであるSARS-CoV-2は、同じくβ-コロナウイルスであり、現在進行中のパンデミックの原因となっている。高病原性コロナウイルスは、人獣共通の保有宿主、主にコウモリ種、において循環し、おそらく中間宿主種を介して、感受性の高いヒト集団に波及数すると考えられている。過去17年間に3件の種を超えた感染事例が発生したことから、類似のウイルスが将来出現する可能性があると示唆される。高病原性コロナウイルスの決定的な特徴は、感染者に重篤な罹患率および死亡率を引き起こす能力であり、大雑把に推定すると、SARS-CoV-2、SARS-CoV-1、およびMERSについて、それぞれ2.3%、9.5%、および34%である(Petrosillo et al、2020年)。一般的に循環しているヒトコロナウイルスは、重篤な罹患率または死亡率を引き起こすことはほとんどない。
病原性コロナウイルスの治療は、主に支持療法である。継続的な努力にもかかわらず、高活性の抗ウイルス薬は存在しない。
本発明は、高病原性コロナウイルス、SARS-CoV-1、MERSおよびSARS-CoV-2、ならびに他の関連する病原性コロナウイルスによって引き起こされる疾患の治療および予防に関する。
コロナウイルスの治療に対する本発明のアプローチは、身体の自然免疫防御を利用する免疫療法である。既存のウイルス感染を排除するための免疫防御のパラダイムは、ウイルス感染細胞を破壊し排除する能力を有する、MHCクラスI拘束性のCD8+細胞傷害性Tリンパ球である。しかしながら、感染性ウイルスに対するT細胞の誘導が、宿主が圧倒される前にその感染を除去できるかどうかは問題である。なぜなら、免疫に対する最大の応答を得るには、典型的には数週間、おそらく数ヶ月を要するからである。したがって、高病原性コロナウイルスの場合、急性感染を制御または排除するために、有効な抗ウイルス性細胞傷害性T細胞応答を迅速に動員するための戦略が必要である。提案されるアプローチは、交差反応性記憶T細胞を刺激することである。記憶T細胞は、ナイーブ細胞よりも刺激に対する応答性が高く、クローン的に非常に急速に増殖することができる。最近の研究では、2019年のウィルスの出現に先立って得られた血液サンプルにおいて、SARS-CoV-2免疫を認識する交差反応性T細胞が同定されている(Grifoni et al、2020年; Weiskopf et al、2020年; Braun et al、2020年; Le et al、2020年)。感冒コロナウイルスであるOC43およびHKU1はSARS-CoV-2とアミノ酸に類似性があるため、これらの交差反応性T細胞は、感冒ヒトコロナウイルスによるそれ以前の感染によって誘導される可能性がある。実際、交差反応性T細胞によって認識されるエピトープをマッピングした最近の研究では、既存の記憶CD4+T細胞が、SARS-CoV-2および感冒コロナウイルスに対して交差反応性であることを実証している(Mateus et al、2020)。交差反応性T細胞の詳細な分子的および構造的基盤は十分には理解されていないが、免疫認識の現在のパラダイムは、SARS-CoV-2および感冒コロナウイルスによって共有されるMHCクラスI拘束性T細胞エピトープ(長さ8~13アミノ酸の連続配列)内の高度に保存されたアミノ酸配列の必要性を予測している。SARS-CoV-2アミノ酸配列の既知の一般的なヒトコロナウイルスの相同領域とのアラインメントでは、推定上のT細胞エピトープにおけるアミノ酸同一性の高い領域は、限られていることを示す。しかしながら、交差反応性が異種ウイルス由来のエピトープ間の実質的なアミノ酸同一性を必要とするという概念は、免疫認識のクローン選択理論に基づいており、この理論では、個々のリンパ球は単一の抗原に特異的、すなわち、1クローン-1特異性であり、交差反応性エピトープ間では密接なアミノ酸同一性が必要であると仮定している。何人かの科学者はこの理論を疑問視しており、T細胞が複数の特異性、1クローン-数百万の特異性を認識すると仮定するT細胞免疫認識の理論を提案している(Mason、1998;Wilson et al、2004;Kersh and Allen、1996;Sewell、2012)。この理論は、ヒトにはわずか1012個しかT細胞が存在せず、ヒトナイーブT細胞プールには<108個の異なるT細胞受容体が存在するが、MHC分子に結合できる20個のアミノ酸のペプチドの理論上の限界は膨大(>1015)であり、翻訳後修飾を含むペプチドを考慮するとさらに大きくなる可能性が高い、という数学的考察に基づいている。T細胞交差反応性に関するこの理論は、高病原性コロナウイルスの治療および予防の両方のための新規なアプローチにつながる。高病原性コロナウイルスに対するT細胞応答と、大多数の個体が過去に曝され、したがって記憶T細胞を有する、1つ又は複数の遺伝的に関連する一般的なヒトコロナウイルスに対するT細胞応答と、を同時に誘導することで、治療と予防という2つのカテゴリーの細胞性免疫を作り出すことができる。この戦略は、交差反応性の部分的に保護的または完全に保護的な記憶T細胞を呼び戻し、高病原性コロナウイルスに特異的なナイーブT細胞を誘導する。これら2つのクラスのT細胞応答の二重作用は、宿主における急性高病原性コロナウイルス感染を迅速に制御および根絶し、重篤な症状および死亡を予防することで治療的であり、同時に、将来の高病原性コロナウイルス感染に対する長期的な防御を提供することで予防的である。
本発明はさらに、病原性コロナウイルスおよび感冒コロナウイルスに対する細胞傷害性Tリンパ球(CD8+)応答、すなわちMHCクラスI拘束性T細胞応答を誘導するための、組成物および方法に関する。
本発明は、パピローマウイルスL1タンパク質のHIループに、システイン残基で挟まれた連続する負電荷アミノ酸が挿入されている、ポリイオンパピローマウイルスのウイルス様粒子(VLP)からなる抗原のためのデリバリープラットフォームに関する。
本発明はさらに、上記VLPと対になる抗原であって、システイン残基の前および/または後に連続する正電荷アミノ酸からなるN末端またはC末端アミノ酸を有する融合ペプチド/タンパク質を含む抗原に関する。以下、この抗原をTAGと称する。本発明はさらに、MHCクラスIエピトープの提示を増強するための、C末端タンパク質プロセシング配列(AAYY)を有するTAGに関する。
特定の実施形態において、本発明は、病原性コロナウイルスに由来する抗原、および、ヒト集団において一般的に循環する遺伝的に最も近縁のヒトコロナウイルスに由来する抗原に関する。本発明はさらに、ウイルス感染細胞におけるウイルスタンパク質の発現量、タンパク質中のMHCクラスI拘束性T細胞エピトープの予測される密度および位置、ならびに、特定のウイルスタンパク質を対象とする細胞傷害性T細胞を同定する経験的研究に基づいた、細胞傷害性T細胞応答を誘導するための抗原の選択に関する。本発明はさらに、ウイルス構造タンパク質およびウイルス非構造タンパク質にそれぞれ由来する、2つの別個の異なるクラスの標的を含む抗原の選択に関する。ウイルス感染細胞において最初に発現されるタンパク質は非構造ウイルスタンパク質であるため、これらのタンパク質を標的とすることにより、ウイルス感染の最も初期の段階で有効な細胞性免疫応答が得られる。本発明はさらに、上記基準、および、追加の基準であって、抗原が病原性コロナウイルスの対応するウイルスタンパク質と少なくとも40%の同一性を共有するウイルスタンパク質の領域を含むという追加の基準に基づく、一般的な非病原性ヒトコロナウイルスの抗原の選択に関する。VLP抗原組成物に関する特定の実施形態において、抗原は、長さ25~30アミノ酸の短いペプチド、長さ約45~55アミノ酸の伸長ペプチド、長さ約110~140アミノ酸の短いタンパク質、または標的抗原の全長アミノ酸配列のいずれかである。
特定の実施形態において、病原性コロナウイルスはSARS-CoV-2であり、遺伝的に関連するヒトコロナウイルスはOC43およびHKU1である。特定の実施形態において、SARS-CoV-2の抗原は、以下のウイルス構造タンパク質:膜タンパク質(M)、ヌクレオカプシドタンパク質(N)、ORF3a、ORF7a、およびスパイク(S)エンベロープタンパク質のS2領域;に由来し、非構造タンパク質の抗原は、nsp6、nsp7、およびnsp12に由来する。特定の実施形態において、OC43およびHKU1の構造タンパク質の抗原は、Mタンパク質およびNタンパク質、ならびにSタンパク質のS2領域に由来し、非構造タンパク質の抗原は、OC43のnsp3、nsp4、nsp6、nsp7およびnsp12タンパク質、ならびに同じウイルスタンパク質由来のOC43に関するHKU1の可変領域に由来する。
本発明はまた、コロナウイルス抗原と対になったポリイオン性パピローマウイルスVLPを調製する方法に関する。本発明はさらに、静脈内、筋肉内、または皮内経路によるVLP抗原組成物の送達方法に関する。特定の実施形態において、VLP抗原組成物は鼻腔内へまたは吸入によって送達され、肺および鼻咽頭組織常在記憶T細胞を刺激し、気道内の部位に優先的に移動する細胞傷害性Tリンパ球を生成する。
本発明は、負に荷電したアミノ酸配列およびL1タンパク質のHIループ中のシステイン残基を含む遺伝子操作されたパピローマウイルスL1ウイルス様粒子(VLP)(以下、ポリイオン性VLPという。)に関する組成物および方法を提供する。本発明はさらに、ヒト、ウシ、ウマ、マウス、ヒツジ、ブタ、シカ、イヌ、ネコ、またはヤギを含む、任意のパピローマウイルス種由来のL1タンパク質に関する。一実施形態において、L1パピローマウイルスタンパク質は、ウシパピローマウイルス1型(BPV1)に由来する(図12)。
特定の実施形態において、遺伝子操作されたBPV1L1タンパク質のHIループは、N末端もしくはC末端または両方の末端にシステイン残基が隣接する、4~10個の連続した負に荷電したアミノ酸を含む。BPV1L1のHIループは、ここではアミノ酸344~357位として定義され、その範囲は14アミノ酸であり、先端は349位のプロリン(P)残基として定義される(図1)。本発明のVLPの負電荷アミノ酸は、グルタミン酸、アスパラギン酸、またはその両方であり得る。様々な実施形態において、5~12個のアミノ酸長さのアミノ酸配列(4~10個の連続した負電荷アミノ酸および1つまたは2つの隣接するシステイン残基)をHIループに挿入することができ、天然アミノ酸を置換しないか、天然アミノ酸のうちの1または複数を置換してもよい。特定の例示的な実施形態において、HIループに挿入されるアミノ酸配列は、8つの負電荷アミノ酸、グルタミン酸またはアスパラギン酸または交互のアスパラギン酸およびグルタミン酸、および、C末端またはN末端システイン残基を含む9残基長であり、ウシパピローマウイルスL1タンパク質のHIループの347~355位にある9つの天然アミノ酸を置換する(表1、挿入物1~4)。他の特定の例示的な実施形態において、HIループに挿入されるアミノ酸配列は、5、6、7、8、10、または11アミノ酸長であり、それぞれ、4、5、6、7、9、または10個のグルタミン酸とC末端システイン残基とを含む。それぞれの挿入物は、ウシパピローマウイルスL1タンパク質のHIループの位置、347~351、347~352、347~353、347~354、346~355、または345~355において、それぞれ、等価な数の天然アミノ酸を置換する(表1挿入物5~10)。特定の例示的な実施形態において、HIループに挿入されるアミノ酸配列は、9残基長であり、8個の負電荷グルタミン酸とC末端およびN末端システイン残基とを含み、ウシパピローマウイルスL1タンパク質のHIループの346~355位の10の天然アミノ酸を置換する(表1、挿入物11)。他の様々な実施形態において、負電荷アミノ酸およびシステインは、HIループに挿入することができ、挿入物を構成する数よりも少ない数の天然アミノ酸を置換することができる。特定の例示的な実施形態において、8個のグルタミン酸およびC末端システイン残基を含む9アミノ酸配列は、ウシパピローマウイルスL1タンパク質のHIループのそれぞれ348~354位、または348~352位、または348~350位においてそれぞれ、7個、または5個、または3個の天然アミノ酸を置換する(表1、挿入物12~14)。他の様々な実施形態において、負電荷アミノ酸およびシステインは、HIループに挿入され、挿入物を含む数よりも最大2つ多い天然アミノ酸を置換することができる。特定の例示的な実施形態において、8つのグルタミン酸およびC末端システイン残基を含む9アミノ酸配列は、ウシパピローマウイルスL1タンパク質のHIループの346~356位の11の天然アミノ酸を置換する(表1、挿入物15)。様々な実施形態において、HIループに挿入されたグルタミン酸-システインアミノ酸配列は、ループの先端と推定される位置においてプロリンを置換してもよく、または、プロリン残基と直後のC末端ロイシン残基との間において天然アミノ酸を何ら除去することなく挿入されてもよい。いくつかの実施形態において、プロリンを置換するかまたはプロリン残基とロイシン残基との間に位置する挿入アミノ酸は、グリシン-セリン-セリン-グリシン(GSSG)リンカーアミノ酸配列によって隣接してもよい。特定の例示的な実施形態において、HIループに挿入されるアミノ酸配列は、GSSGアミノ酸が隣接しているか隣接していない、8つのグルタミン酸およびC末端システイン残基を含み、349位のプロリンを置換するか、またはウシパピローマウイルスL1タンパク質のHIループのアミノ酸349位と350位との間に位置する(表1、挿入物16~19)。
本発明はさらに、VLPに連結させる抗原に関し、この抗原は、システイン残基が先行するおよび/または後続する4~10の連続した正電荷アミノ酸からなるN末端またはC末端アミノ酸を有する融合ペプチド/タンパク質を含む。この抗原を以後、TAGと称する。TAGは、静電相互作用と、VLP上のシステイン残基とTAG中のシステイン残基との間の酸化還元反応と、の複合作用によって、抗原をポリイオン性パピローマウイルスVLPに結合させることができる。本発明はさらに、MHCクラスIエピトープの提示を増強するためのC末端タンパク質プロセシング(分解処理)配列(AAYY)を有するTAGに関する。TAGが抗原のC末端に付加される場合、タンパク質プロセシング配列(AAYY)は、ペプチド/タンパク質抗原のN末端に配置される。
特定の例示的な実施形態において、TAGは、正電荷を有する、アミノ酸、アルギニン(R)、リジン(K)、またはヒスチジン(H)の群を含み、配列は8アミノ酸長である(表2、Tag-1、-2、-3)。他の特定の例示的な実施形態において、TAGは、8アミノ酸長のRKHRKHRKの反復モチーフを含む(表2、TAG-4)。他の特定の例示的な実施形態において、TAGは、4、5、6、または7つの連続したアルギニンにシステイン残基が続くアミノ酸配列(表2、TAG-5、-6、-7、および-8)、または、9または10個のアルギニンにシステイン残基が続くアミノ酸配列(表2、TAG-9および-10)を含む。他の特定の例示的な実施形態において、TAGは、システイン残基が先行するか、またはシステイン残基によってN末端およびC末端に隣接する、8個の正アルギニンからなる(表2、TAG-11および-12)。
本発明はさらに、細胞傷害性(MHCクラスI拘束性)T細胞応答を誘導する目的のための、ポリイオン性パピローマウイルスVLPおよびTAG配列を有する標的抗原の組成物に関する。特定の実施形態において、本発明は、感冒コロナウイルスおよび病原性コロナウイルスに対する細胞傷害性T細胞応答を誘導する目的で、ポリイオン性VLPに連結されたコロナウイルス抗原に関する。
ウイルスのプロテオーム全体がT細胞応答の標的となり得る。しかしながら、構造タンパク質は、感染細胞において最も豊富に発現するウイルスタンパク質であり、したがってMHC分子への効率的な提示を可能にするため、抗原特異的T細胞応答のための特に有効なターゲットである。コロナウイルスに対する細胞性免疫応答の経験的研究も、細胞性免疫応答の主要な標的としての構造タンパク質の重要性を支持している(Liu et al、2017の概説)。細胞傷害性T細胞応答は、MHCクラスI拘束性T細胞エピトープに向けられる。これらのエピトープは、最も一般的には9アミノ酸長であり、8~13アミノ酸長の範囲であり得る。ヒトおよび他の哺乳動物におけるMHCクラスI対立遺伝子の遺伝的不均一性により、ウイルスタンパク質の中に多数のエピトープが存在する可能性がある。可能性のある非常に多くのエピトープを網羅するために、抗原は、標的タンパク質の全アミノ酸配列であるか、または全アミノ酸配列を一緒に含む抗原の短い断片の集合である必要がある。ポリイオン性VLPSに連結された抗原断片の好ましい長さは、代替形態抗原提示経路を介したT細胞応答の誘導のためのエピトープ提示効率の経験的関数である。加えて、製造上の考慮が、抗原の好ましい長さの選択に影響を及ぼし得る。
SARS-CoV-2に対する細胞傷害性T細胞応答を誘導するためのポリイオン性パピローマウイルスVLP組成物に関する好ましい実施形態は、膜(M)、ヌクレオカプシド(N)、スパイク(S)のS2領域、ORF3a、およびORF7a構造タンパク質、ならびにnsp6、nsp7、およびnsp12非構造タンパク質に由来の抗原を含む。
Mタンパク質は222アミノ酸長であり、抗原領域はaa6~221に及ぶ。Nタンパク質は、419アミノ酸長である。MHCクラスI拘束性T細胞エピトープに関する経験的研究と予測アルゴリズムに基づくと、抗原領域はaa51~369である。スパイク(S)タンパク質は1270アミノ酸長であり、受容体結合ドメイン(aa330-583)を含むS1(aa1-661)とS2(aa662-1270)との2つの領域を含む。T細胞エピトープは、Sタンパク質にわたって広く分布している。S1領域は、ビリオン表面に露出したアミノ酸の大部分を含み、中和抗体を含む体液性免疫応答の主要な標的である。この領域は、抗体依存性増強(ADE)を媒介する抗体の誘導のような、有害な作用を起こし得る抗体応答の意図しない誘導を回避するために、細胞性免疫の誘導を意図したワクチンから除外することが好ましい。ORF3aは275アミノ酸長である。経験的研究によると、T細胞応答のための標的抗原としてORF3aを選択することが支持される。予測アルゴリズム(MHC-NP:Sebastien Giguere, Alexandre Drouin, Alexandre Lacoste, Mario Marchand, Jacques Corbeil,およびFrancois Lavioletteによって開発されたMHCによる、自然にプロセシングされたペプチドの予測;http://tools.iedb.org/mhcnp/)によると、5つの代表的な対立遺伝子について、C末端はN末端と比較して、推定MHCクラスI制限エピトープの密度が高いことが示される(N末端104アミノ酸にわたる20個に対して、C末端171アミノ酸にわたる67個)。経験的研究は、細胞傷害性T細胞応答の標的として、121アミノ酸のタンパク質であるORF7aを選択することを支持している。SARS-CoV-2のORF1abは、7096アミノ酸のポリタンパク質である。このポリタンパク質は、ウイルス感染細胞内でタンパク質分解的に処理(プロセシング)されて、ウイルスのライフサイクルにおいて多様な機能を果たす複数の非構造タンパク質を生成する。タンパク質はウイルス感染細胞中に低存在量で存在し、このため、細胞性免疫応答の主要なターゲットではないが、最近の研究で、いくつかのnspタンパク質に対する細胞傷害性T細胞応答が、SARS-CoV-2感染患者の血液中で、また、COVID19前に何人かの健康な血液ドナーから得られた血液中で、検出され得ることが示された(Grifoni et al、2020; Le et al、2020)。これらのタンパク質は、感染細胞において最初に発現するため、細胞傷害性T細胞にとって魅力的な標的である。これらのウイルス感染細胞を迅速に死滅させることにより、細胞が感染性ウイルスを産生するのを防ぐことができる。抗原特異的T細胞の頻度および応答する対象の割合からみると、CD8+T細胞応答の主要な標的は、nsp6、nsp7、およびnsp12タンパク質である。
ポリイオン性VLPに連結された抗原のアミノ酸長は、8~14アミノ酸の定義されたエピトープまたはそれより長いアミノ酸配列から、ウイルスタンパク質またはいくつかのウイルスタンパク質の融合体の抗原領域の全長アミノ酸配列まで、の範囲とすることができる。例示的な実施形態において、抗原は、長さが約25~30アミノ酸の短いペプチド、長さが約45~55アミノ酸の伸長ペプチド、長さが約110~140アミノ酸の短いタンパク質、または標的とするウイルスタンパク質の抗原領域の全長アミノ酸配列である。抗原領域の全長アミノ酸配列よりも短い抗原断片の場合、特定の実施形態は、標的ウイルスタンパク質の抗原領域の全アミノ酸を含む抗原のセット(集合)を含む。SARS-CoV-2のMタンパク質に対する抗原を、抗原領域の、短いペプチド、伸長ペプチド、短いタンパク質、または全アミノ酸配列として具体化し、表3に示す。SARS-CoV-2のNタンパク質に対して同等の抗原を表4に示す。SARS-CoV-2のS2、ORF3a、およびORF7a構造タンパク質について、同等に設計された抗原を表5~7に示す。非構造タンパク質nsp6およびnsp7の抗原を表8に示し、nsp12タンパク質に対する抗原を表9および表10に示す。短いペプチドと伸長ペプチドは10アミノ酸が重なり、短いタンパク質抗原は11アミノ酸が重なっている。最終製剤において、ペプチド、タンパク質、または全長抗原は、上記のようにTAGを有する。
感冒コロナウイルスへの事前の曝露によって誘導される記憶T細胞を刺激するためのポリイオン性パピローマウイルスVLP組成物に関する好ましい実施形態は、構造タンパク質と非構造タンパク質の両方に由来する抗原を含む。T細胞認識の複数の特異性理論は、T細胞の交差反応性の構造的基礎を分子レベルで定義していない。本発明者らは、本明細書において、交差反応性T細胞エピトープを含有するウイルスタンパク質またはウイルスタンパク質のサブ領域を、一般的な感冒コロナウイルスと特定の病原性コロナウイルスの相同アミノ酸配列にわたって40%を超える(>40%)平均アミノ酸同一性を有するものと定義する。
特定の実施形態において、共通のコロナウイルスはOC43およびHKU1であり、交差反応性T細胞応答を誘導するための抗原は、M、N、およびS2構造タンパク質、ならびにnsp3、nsp4、nsp6、nsp7およびnsp12非構造タンパク質、に由来する。OC43およびHKU1のアミノ酸配列が、標的抗原の相同アミノ酸間で平均して40~80%超で(>40-80%)共有する場合、OC43のアミノ酸配列のみが標的抗原として使用される。他の特定の実施形態において、HKU1のアミノ酸配列は、抗原である。
OC43のMタンパク質は長さが230aaであり、aa14-226の領域内でSARS-CoV-2のMタンパク質と40.8%の同一性を共有する(図2)。OC43のMタンパク質とHKU1のMタンパク質とのアラインメントは、有意な変動性の領域(アミノ酸同一性<80%)を示さない(図3)。OC43のNタンパク質は長さが448aaであり、SARS-CoV-2のNタンパク質と平均で36%のアミノ酸同一性を有する。非整列領域aa266~269、aa341~349、およびaa382~387を除く、aa99~400由来のOC43の抗原領域は、OC43のNタンパク質のアミノ酸配列と整列しないaa221~224を除いた後、SARS-CoV-2のNタンパク質の相同領域と43.5%の同一性を有する(図4A)。aa64~88位のOC43のNタンパク質のN末端アミノ酸は、SARS-CoV-2のNの対応する領域と52%の同一性を有する(図4B)。OC43のNタンパク質とHKU1のNタンパク質とのアラインメントは、2つのウイルスアミノ酸配列間のアミノ酸同一性が33%~50%の範囲と低いレベルの3つの抗原領域を示す。(図5A、5B、および5C)。OC43のSタンパク質は1353アミノ酸であり、S1とS2の2つのサブ領域から構成される。S1はaa1-789まで伸びており、aa70-552間の整列領域においてSARS-CoV-2のS1領域と低いアミノ酸同一性(24%)を示す。対照的に、ウイルスのS2領域は、aa898-1153およびaa1228-1302の2つの抗原領域を含み、これらは、それぞれ平均で52.72%および51.3%のアミノ酸同一性を有する(図6)。HKU1の相同領域とOC43のaa898-1153とのアラインメントは、変動性の高い領域を含むことなく83.5%という全体的な同一性を示し、aa1228-1303間のOC43の抗原領域は、HKU1の相同S2領域と70.7%の同一性を有する(図7)。
例示的な実施形態において、抗原は、長さが約25~30アミノ酸の短いペプチド、長さが約45~55アミノ酸の伸長ペプチド、長さが約110~140アミノ酸の短いタンパク質、または標的ウイルスタンパク質の抗原領域の全長アミノ酸配列である。抗原領域の全長アミノ酸配列よりも短い抗原断片に関し、特定の実施形態は、標的ウイルスタンパク質の抗原領域の全アミノ酸を含む抗原のセットを含む。OC43のNタンパク質およびHKU1の可変領域についての同等の抗原を、抗原領域の、短いペプチド、伸長ペプチド、短いタンパク質、または全アミノ酸配列として具体化して、表11に示す。OC43のS2タンパク質およびHKU1の可変領域について同等の抗原を表12に示し、OC43のS2タンパク質およびHKU1の可変領域に対する抗原を表13に示す。短いペプチドと伸長ペプチドは10アミノ酸が重なり、短いタンパク質抗原は11アミノ酸が重複する。最終製剤において、ペプチド、タンパク質または全長アミノ酸配列には、上記のようにTAGが含まれる。
OC43コロナウイルスのORF1abは、7095アミノ酸からなるポリタンパク質をコードしている。このポリタンパク質は、ウイルス感染細胞内でタンパク質分解的に処理されて、ウイルスのライフサイクルにおいて多様な機能を果たす複数の非構造タンパク質を生成する。抗原特異的T細胞の出現頻度および応答する対象の割合から、CD8+T細胞応答の主要な標的は、nsp3、nsp4、nsp6、nsp7、およびnsp12タンパク質である。これらの非構造タンパク質からの抗原の選択はまた、感冒ウイルスとSARS-CoV-2との間のアミノ酸配列が平均して>40%の同一性であるという、追加的な考慮に基づく。
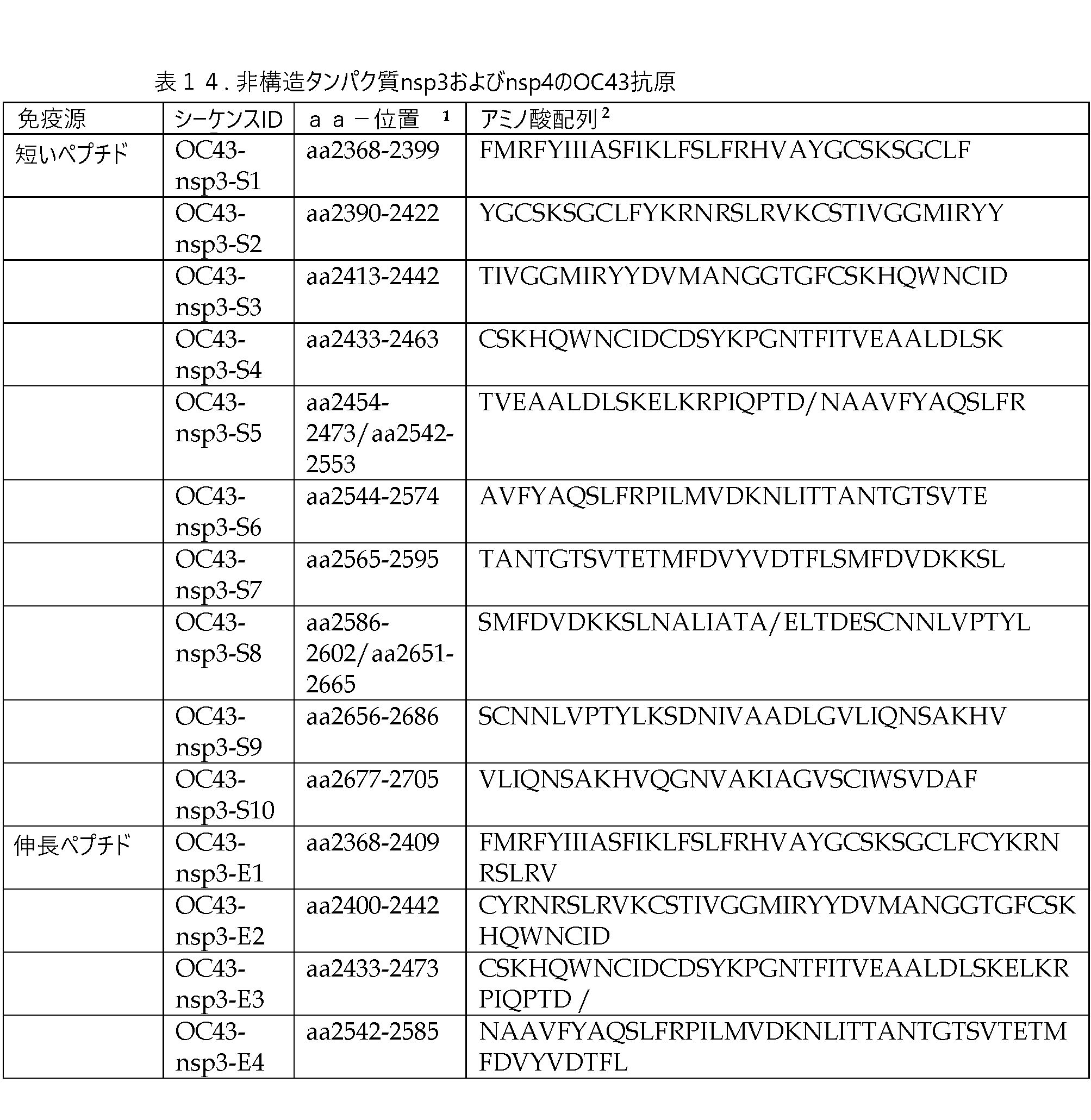
nsp3タンパク質は、1,945アミノ酸長である。SARS-CoV-2とOC43のnsp3アミノ酸配列との間の全体的な同一性は26%である。しかし、384アミノ酸のC末端領域は39%の同一性を有し、その中に、OC43のアミノ酸配列とSARS-CoV-2の相同アミノ酸配列との間で52.8%、41%、および43.6%という同一性を有する連続したアミノ酸からなる3つの領域を含む(図8A~C)。nsp4タンパク質は長さが500個のアミノ酸であり、SARS-CoV‐2とOC43のnsp4タンパク質は、42%のアミノ酸同一性を有する。SARS-CoV-2とOC43の相同アミノ酸配列間では、nsp4タンパク質の中央部分またはC末端における3つのサブ領域は、それぞれ45、133および50アミノ酸長であり、それぞれ53.3%、47.4%および72%のアミノ酸同一性を有する(図9A~C)。nsp6タンパク質は290アミノ酸長であり、SARS-CoV-2およびOC43の相同アミノ酸配列は30.5%の同一性を有する。C末端の84アミノ酸は54.8%の同一性を有し、N末端の31アミノ酸領域は41.9%の同一性を有する(図10A~B)。nsp7タンパク質は83アミノ酸長であり、SARS-CoV-2とOC43のnsp7のアミノ酸配列は46%の同一性を有する。SARS-CoV-2のC末端13アミノ酸を除くと、SARS-CoV-2とOC43のnsp7タンパク質のアミノ酸同一性は、55.2%である(図11)。nsp12タンパク質は、932アミノ酸長である。SARS-CoV-2感染患者における抗原特異的細胞性免疫応答の経験的研究では、T細胞エピトープの大部分が、タンパク質のaa125~375およびaa520~920に包含される領域に位置づけられた(Grifoni et al、2021)。OC43のORF1ab内の2つのアミノ酸配列は、SARS-CoV-2のnsp12タンパク質のaa125~275断片内の対応するアミノ酸と、61.5%および67%の同一性を有し(図13A~B)、OC43のORF1abの341aaは、SARS-CoV-2のnsp12タンパク質のaa520~920断片内の対応するアミノ酸と、76.8%の同一性を有する(図13C)。OC43とHKU1の非構造タンパク質は、~90%の同一性を有し、アミノ酸可変性の拡張領域は有さない(同一性<80%)。
例示的な実施形態において、抗原は、長さが約25~30アミノ酸の短いペプチド、長さが約45~55アミノ酸の伸長ペプチド、長さが約110~140アミノ酸の短いタンパク質、または標的ウイルスタンパク質の抗原領域の全長アミノ酸配列である。抗原領域の全長アミノ酸配列よりも短い抗原断片について、特定の実施形態は、標的ウイルスタンパク質の抗原領域の全アミノ酸を含む抗原のセットを含む。
nsp3およびnsp4タンパク質に対する抗原を、抗原領域の、短いペプチド、伸長ペプチド、短いタンパク質、または全アミノ酸配列として具体化して、表14に示す。OC43のnsp6タンパク質およびnsp7タンパク質について同等に設計された抗原を表15に示し、nsp12については表16および17に示す。短いペプチドと伸長ペプチドは10アミノ酸が重なり、短いタンパク質抗原は11アミノ酸が重複する。最終製剤において、ペプチド、タンパク質または全長アミノ酸配列には、上記のTAGが含まれる。
nsp3およびnsp4タンパク質に対する抗原を、抗原領域の、短いペプチド、伸長ペプチド、短いタンパク質、または全アミノ酸配列として具体化して、表14に示す。OC43のnsp6タンパク質およびnsp7タンパク質について同等に設計された抗原を表15に示し、nsp12については表16および17に示す。短いペプチドと伸長ペプチドは10アミノ酸が重なり、短いタンパク質抗原は11アミノ酸が重複する。最終製剤において、ペプチド、タンパク質または全長アミノ酸配列には、上記のTAGが含まれる。
実施例1:遺伝子操作されたBPVのL1遺伝子を有する組換えバキュロウイルスの生成
Kozakコンセンサスを有し、各末端に特有の制限部位(EcoR1/Not1)を持つBPVのL1のオープンリーディングフレーム(ORF)全体を、PCRベースの遺伝子合成によって人工的に操作し、pUC18ベクターにクローニングした。昆虫細胞で効率的に発現させるために、ORF全体を、キイロショウジョウバエの好ましいコドンを用いてコドン修飾した。ORFは、アスパラギン酸残基またはグルタミン酸残基と、システイン残基と、を有するペプチドの挿入物を含み、表1に記載されるように、HIループに挿入され、挿入物-1~挿入物-19と命名する。
修飾BPV L1遺伝子は、pORBバキュロウイルス転移ベクターのEcoR1/Not1部位間にサブクローニングされる。この転移ベクターを、製造業者によって提案されている好ましい市販のトランスフェクション試薬を用い、線状バキュロウイルスDNAとともに、ヨトウガのsf9細胞中にコトランスフェクションした。トランスフェクションから5日後、回収した組換えバキュロウイルスを、sf9細胞の大量感染させてさらに増幅させた。6ウェルプレート中で増殖しているイラクサキンウワバの2×106のHigh Five細胞を用い、バキュロウイルスストックの連続希釈物で感染させることで、修飾L1タンパク質の発現を確認するための小規模感染を行った。感染の72時間後、細胞を500μlのRIP A緩衝液中で溶解し、清澄化した溶解液をSDS-PAGE分析に供して、予測される分子量55kDaのタンパク質の過剰発現を検出した。
実施例2:組換えバキュロウイルスからのポリイオン性VLPの産生
VLPの産生のために、スピナーフラスコ中で増殖している約2×109のイラクサキンウワバのHigh Five細胞を、500mlのTNM-FH/10%FBS中で、所定量の高力価組換えバキュロウイルスストックに感染させた。27℃で96時間培養した後、細胞を回収し、2,000rpm(Sorvall,FH18/250遠心機)で5分間の遠心分離した。細胞ペレットを、プロテアーゼ阻害剤(Roche Complete ULTRA、10mlあたり1錠)を含む抽出緩衝液(20mMリン酸緩衝液、pH6.5、1M NaCl、0.1mM CaCl2、50μm FeCl2)中に再懸濁し、37℃での融解と-80℃エタノールバスでの凍結とを5サイクル実施した。溶解液を8000rpmで1時間回転させ、バキュロウイルス粒子を除去した。清澄化された溶解液を、等容量のバートレルDF(Vertrel DF、Fisher Scientific製)で10分間抽出した。水層を、40%ショ糖クッション上に重層し、SW32Tiローターで、32,000rpm、1.5時間で遠心分離した。ショ糖ペレットを20mMリン酸塩pH8.0、0.5M NaCl、5mM MgCl2に再懸濁し、250U/ml塩活性ヌクレアーゼ(Arcticzymes製)と共に37℃で30分間培養した。20mMリン酸塩pH6.5、0.5M NaClで透析した後、VLP液を、0.01%Tween80、0.05%カルボキシメチルセルロース、50μM FeCl2に調整し、4℃で保存した。純度をSDS-PAGEゲル分析によって評価し、タンパク質濃度をブラッドフォード色素法およびUV分光法によって測定した。VLPの直接可視化を容易にするために、希釈粒子のアリコートを、300メッシュのformvar/炭素被覆銅グリッド上に置き、2%リンタングステン酸(pH=7.0)でネガティブ染色し、透過型電子顕微鏡(TEM)で観察した。表1から挿入物を有するキメラVLPは、より小さなカプソメア様構造からの形成と一致し、天然のHPV1型VLPに類似する多彩な外観を備えた、直径~50nmのカプシド様構造を生成することが示された。
実施例3:抗原のポリイオン性VLPへのコンジュゲーション
VLPへのコンジュゲーションのために、ペプチドを5mg/mlで蒸留水に溶解させた(水に溶けない場合、DMSOに5mg/mlで溶解)。好ましい濃度2.5mg/mlであるが、緩衝液中の溶解性が低い場合にはより低い濃度のペプチドを、10mMのBond-breaker TCEP溶液(Thermo Fisher Scientific製)を用いて50℃で20分間還元した。20mMリン酸緩衝液(pH6.5、0.15M NaCl)中で透析した後、VLPタンパク質(1mg/ml)とペプチドを、溶解特性に基づきペプチド:L1タンパク質のモル比を4:1~16:1として、4mMグルタチオンジスルフィド(GSSG)および0.8mM還元グルタチオン(GSH)の存在下で混合し、37℃で一晩培養した。未反応のペプチドを除去するために、100万kDaのカットオフの透析チューブを使用して、反応物を20mMリン酸、pH6.5、0.5M NaClに対して透析した。VLP-ペプチド溶液は、0.01%Tween80、0.05%カルボキシメチルセルロース、0.5mM GSSG、および0.05mM GSHに調整し、アリコートした後、-20℃で保存した。VLPに結合したペプチドの量を、SDS-PAGE分析と、既知量のペプチド標準曲線からのサンプル-ペプチドバンド密度の内挿によって決定した。ゲルを、BioRad製のChemiDocXR撮影装置でスキャンし、画像をNIHのImageJソフトウェアで解析した。
実施例4:細胞溶解CD8+T細胞応答を誘導するための、HIループに様々な挿入物を有するVLPの例示的な実施形態の能力
L1タンパク質(表1)中の種々例示的な挿入物を用いて生成されたVLPは、TAGを介して、C57BL/6マウスの遺伝的バックグラウンドにおいて認識されるMHCクラスI拘束性エピトープをコードする代表的なペプチドに連結した。マウスは、以下に記載されるように、皮内経路によって免疫した。免疫応答は、以下に記載するように測定した。代表的な抗原に連結された複数の挿入物を用いて製剤化したVLPは、抗原特異的インターフェロン-γ分泌CD8+T細胞(t検定比較による平均応答に有意差なし)の細胞傷害性T細胞応答を、同程度の頻度で誘導することが示された。
実施例5:例示的実施形態のTAGによって抗原に連結されたVLPの細胞傷害性CD8+T細胞応答を誘導する能力
C57BL/6マウスの遺伝的バックグラウンドにおいて認識されるMHCクラスI拘束性エピトープをコードする代表的なペプチドを、表2に列挙されるTAGを用いて、>90%の純度で化学的に合成する。TAG-1~-4、-11および-12を有するペプチドが、挿入物-1を有するポリイオン性VLPに連結される。TAG5~10を有するペプチドは、それぞれ挿入物5~10を有するVLPに連結される。マウスは以下に記載されるように、皮内経路によって免疫される。免疫応答は、以下に記載するように測定される。複数のTAGを用いて製剤化された代表的な抗原に連結されたVLPは、抗原特異的なインターフェロン-γ分泌CD8+T細胞を同程度の頻度で誘導することが示された(t検定比較による平均応答に有意差なし)。
実施例6:ポリイオン性VLPによる免疫化と、抗原特異的CD8+T細胞応答の検出
6~8週齢のC57BL/6Jマウスに、1週間隔で3回、それぞれ5~50ugのVLP-ペプチドを皮内注射することによって免疫した。コントロールとして、C57BL/6Jマウスを、非標識(ペプチドなし)VLPタンパク質で免疫する。VLP-ペプチド免疫原は、表3~12に記載されている、異なるSARS-CoV-2抗原およびOC43抗原のための伸長ペプチドのセットを使用して製剤化された。皮内免疫化の場合、VLP/抗原ワクチンを、剃毛したマウスの背中の皮膚に、単回または分割用量で注射した。
免疫学的アッセイ用の試料を提供するために、ワクチンの最終投与から10~14日後にマウスを屠殺し、脾臓を解剖した。脾細胞(または肺組織由来のCD3+細胞の懸濁液)を、ブレフェルジンA(10μg/ml)の存在下、標的抗原の全長アミノ酸配列にわたる長さ11アミノ酸(8アミノ酸が重複)の重複ペプチドのプール1ug/mlを用い、5%CO2中37℃で一晩刺激した。細胞を、Zombie green(商標)固定可能な生存率解析用色素で染色し、固定緩衝液で処理し、Cytolast中で保存した。細胞は、透過処理緩衝液で透過処理し、Brilliant Violet BV(商標)510標識抗マウスCD3、クローン17A2、PerCPCy5.5標識抗マウスCD8α、クローン53-6.7、およびPE標識抗マウスIFNγ、クローンXMG1.2で、染色した。試薬は、市販のものを使用した。LSR-IIまたは同等のフローサイトメーターでフローサイトメトリーを行い、FACSDivaソフトウェアまたはFlowJoソフトウェアを用いてデータを解析した。前方散乱と側方散乱のパラメータでゲーティングを行い、リンパ球と単一細胞を選択した。死細胞の除去後、CD8+Tリンパ球を、ゲーティングされたリンパ球のCD3/CD8ドットプロット上で同定し、インターフェロン-γ(IFNγ)分泌細胞を、ゲーティングされたCD8+T細胞のCD8/IFNγドットプロット上で同定した。最低30,000個のCD8+T細胞を分析する。SARS-CoV-2のM、N、S2、ORF3a、ORF7a、nsp6、nsp7、およびnsp12抗原のVLPペプチド免疫原、ならびにOC43のM、N、S2、nsp6、nsp7、およびnsp12抗原の免疫原は、検出可能な抗原特異的なインターフェロン-γ分泌CD8+T細胞を誘導することが示された。
実施例7:チャレンジモデルにおけるSARS-CoV-2疾患からマウスを保護するポリイオン性VLP SARS-CoV-2ワクチンの能力
マウス株:C57BL/6系マウスにおいて、内因性マウスACE2(mACE2)をヒトACE2に置換するために、CRISPR/Cas9ノックイン技術を用いて作製した安定ヒト化アンギオテンシン変換酵素II(ACE2)マウスを、SARS-CoV-2チャレンジに使用した(Sun et al.、2020年)。
マウス免疫化:HIループ中に例示的な挿入物を有するVLPを、特定の挿入物に対して例示的かつ適切なTAGを有する例示的な伸長ペプチドのセットと共に製剤化した。ポリイオン性VLPワクチンを、SARS-CoV-2抗原のM、N、S2、ORF3a、ORF7a、nsp6、nsp7および/またはnsp12のSARS-CoV-2抗原と共に製剤化した。ペプチドは、シーケンスIDが伸長を表す-E1、-E2、-E3などで終わる表3~10のリストから選択される。ペプチドの正確な数は、表に記載されるように抗原によって異なる。好ましい挿入物は、天然アミノ酸の置換を伴う、ウシパピローマウイルス1型L1タンパク質のHIループにおけるアミノ酸位置347~355のE8Cアミノ酸配列であり(挿入物-1、表1)、好ましいTAGは、CRRRRRRRRCAAYYである(TAG-1、表2)。1または複数のSARS-CoV-2抗原に対するペプチドのセットを有する、挿入物およびTAGの追加の組合せも、他の可能な実験から情報を得て試験される。VLP/抗原構築物は、上記のように作製される。マウスは、皮内経路、鼻腔内/肺経路、または両方の経路で、同時に免疫される。皮内免疫の場合、VLP/抗原ワクチンを、剃毛したマウスの背中の皮膚に、単回または分割用量で注射する。鼻腔/肺免疫の場合、VLP/抗原構築物を、軽く麻酔したマウスの鼻に滴下投与(10μl)する。麻酔をかけたマウスでは、鼻腔内に投与されたワクチンもマウスに吸入されるため、肺組織と鼻咽頭組織の両方に送達される。
マウスチャレンジ:鼻腔内感染のために、高齢(30週齢)hACE2マウスを、精密気化器を用いて送達されるイソフルランで麻酔し、次いで、4×105pfuのSARSCoV-2で鼻腔内感染させる。その後、マウスの体重を測定して毎日モニターし、感染後6日目に、血清採取と組織処理のために屠殺する。脾臓と肺組織を、ウイルスRNA量の分析、組織病理学、および細胞性免疫応答の測定のために採取する。
ウイルスRNA量の測定:肺組織中のウイルスRNAを、プロトコールに従ってRNeasyMiniキット(QIAGEN)を用いて抽出する。ウイルスRNAの定量化は、SARS-CoV-2のS遺伝子を標的とするRT-qPCRによって行う。RT-qPCRは、以下のプライマーおよびプローブ:CoV-F3、CoV-R3およびCoV-P3(Sun et al、2020)で、PrimeScript(商標)1ステップRT-PCR用キット(タカラ)を用いてを行う。
抗原特異的CD8+T細胞応答:脾臓細胞と、肺ホモジネートから回収したCD3+T細胞と、についてのCD8+T細胞応答を、上記のように測定する。
SARS-CoV-2ポリイオン性VLPワクチンは、標的抗原(M、N、S2、ORF3aおよびORF7a)のそれぞれに対して、脾細胞および肺組織由来のCD3+T細胞において検出可能な抗原特異的CD8+T細胞応答を誘導することが示された。さらにワクチン接種されたマウスは、模擬ワクチンを接種されたマウスと比較して、ウイルスRNA量が優位に低減され、肺病理学的所見が少ないことが示された。ワクチンはまた、肺におけるウイルスRNAの発現を減少させることも示された。なお、ワクチンが滅菌免疫提示することは期待されない。
実施例8:チャレンジモデルにおいてシリアンハムスターをSARS-CoV-2感染から保護するポリイオン性VLPのSARS-CoV-2ワクチンの能力
動物種:シリアンハムスターは、SARS-CoV-2感染に対して非常に感受性が高く、ワクチンの防御効果を評価するのに最適な小動物モデルである(Rosenke et al、2020;Chan et al.、2020)。6~8週齢のシリアンハムスター(Mesocricetus auratus)を、Jackson Laboratoriesから購入した。
ハムスター免疫化:HIループ中に例示的な挿入物を有するVLPを、特定の挿入物に対して例示的かつ適切なTAGを有する例示的な伸長ペプチドのセットと共に製剤化した。ポリイオン性VLPワクチンは、M、N、S2、ORF3a、ORF7a、nsp6、nsp7および/またはnsp12のSARS-CoV-2抗原と共に製剤化される。ペプチドは、シーケンスIDが伸長を表す-E1、-E2、-E3などで終わる、表3~10のリストから選択される。ペプチドの正確な数は、表に記載されるように抗原によって異なり、-E伸長指定を有する全ての抗原またはより少ない抗原を含み得る。好ましい挿入物は、天然アミノ酸の置換を伴う、ウシパピローマウイルス1型L1タンパク質のHIループのアミノ酸位置347~355におけるE8Cアミノ酸配列であり(挿入物-1、表1)、好ましいTAGは、CRRRRRRRRCAAYY(TAG-1、表2)である。1または複数のSARS-CoV-2抗原に対するペプチドのセットを有する、追加の挿入物およびTAGの組合せも、他の可能な実験から情報を得て試験することができる。VLP/抗原構築物を上記のように作製する。ハムスターは、皮内経路、鼻腔内経路、または両方の経路を、単独で、または組み合わせて、免疫される。皮内免疫は、上記のように行う。鼻腔内免疫の場合、VLP/抗原構築物を、軽く麻酔したマウスの鼻に滴下(10μl)して投与する。
ハムスターチャレンジ:自然の感染経路を模擬するために、ワクチン接種された動物と対象動物(接触動物)を同じケージに共同収容することによって、以前に感染した動物(指標動物)に曝露する。指標動物に対する鼻腔内感染のために、ハムスターを、精密気化器を用いて送達されるイソフルランで麻酔し、次いで、100組織培養容量50(TCID50)のSARSCoV-2ウイルスで鼻腔内感染させる。CDCからのSARS-CoV-2単離株nCoV-WA1-2020(MN985325.1)、または適切な代替単離株を入手し、VeroE6細胞中で増殖させる。TCID50用量は、VeroE6細胞中のウイルスストックの滴定によって決定される。感染したハムスター、および同時収容されたワクチン接種ハムスターおよび対照のナイーブハムスターの体重を測定し、疾患の臨床徴候がないか毎日モニターした。RT-qPCRによる感染をモニターするために、軽く麻酔したナイーブ接触動物(ワクチン接種および対照)および指標動物(以前に感染させた)に、150ulのPBS/0.3%BSAを両鼻孔に滴下して回収することで、鼻洗浄液を10日間毎日回収した。
ウイルスRNA量の測定:肺組織中のウイルスRNAを、プロトコールに従ってRNeasyMiniキット(QIAGEN製)で抽出した。ウイルスRNAの定量は、SARS-CoV-2のS遺伝子を標的とするRT-qPCRによって行った。RT-qPCRは、以下のプライマーとプローブ:CoV-F3、CoV-R3およびCoV-P3(Sun et al.、2020)で、PrimeScript(商標)1ステップRT-PCR用キット(タカラ)を用いてを行った。
SARS-CoV-2ポリイオン性VLPワクチン接種ハムスターは、感染した指標ハムスターに曝露した後、偽ワクチン接種マウスよりも、鼻洗浄液中のウイルスRNA量が有意に減少することが示された。
[表]









































Braun,J., Loyal,L., Frentsch,M., Wendisch,D., Georg,P., Kurth,F., Hippenstiel,S., Dingeldey,M., Kruse,B., Fauchere,F., Baysal,E., Mangold,M., Henze,L., Lauster,R., Mall,M.A., Beyer,K., Rohmel,J., Voigt,S., Schmitz,J., Miltenyi,S., Demuth,I., Muller,M.A., Hocke,A., Witzenrath,M., Suttorp,N., Kern,F., Reimer,U., Wenschuh,H., Drosten,C., Corman,V.M., Giesecke-Thiel,C., Sander,L.E., and Thiel,A. (2020). SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19(健常ドナーおよびCOVID-19罹患者におけるSARS-CoV-2反応性T細胞). Nature.
Chan,J., Zhang,A.J., Yuan,S., Poon,V., Chan,C., Lee,A., Chan,W.M., Fan,Z., Tsoi,H-W., Wen,L., Liang,R., Cao,J., Chen,Y., Tang,K., Luo,C., Cai,J-P., Kok,K-H., Chu,H., Chan,K-H., Sridhar,S., Chen,Z. Chen,H., To,K.K-W., Yuen,K-Y. (2020). Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility(ゴールデンシリアンハムスターモデルにおけるコロナウイルス疾患2019(COVID-19)の臨床的および病理学的症状のシミュレーション:疾患の病因および伝染性の意味). Clin Infect Dis. 71:2428-2446.
Chen,Y., Liu,Q., and Guo,D. (2020). Emerging coronaviruses: Genome structure, replication, and pathogenesis(新型コロナウイルス:ゲノム構造、複製、および病因). J Med. Virol.
Grifoni,A., Weiskopf,D., Ramirez,S.I., Mateus,J., Dan,J.M., Moderbacher,C.R., Rawlings,S.A., Sutherland,A., Premkumar,L., Jadi,R.S., Marrama,D., de Silva,A.M., Frazier,A., Carlin,A.F., Greenbaum,J.A., Peters,B., Krammer,F., Smith,D.M., Crotty,S., and Sette,A. (2020). Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals(COVID-19感染者と非感染者における、SARS-CoV-2コロナウイルスに対するT細胞応答の標的). Cell 181, 1489-1501.
Grifoni,A., Sidney,J., Vita,R., Peters,B., Crotty,S., Weiskopt,D., and Sette,A. (2021). SARS-CoV-2 human T cell epitopes: Adaptive immune response against COVID-19(SARS-CoV-2ヒトT細胞エピトープ:COVID-19に対する適応免疫応答). Cell Host & Microbe 29, 1076-1092.
Kersh,G.J. and Allen,P.M. (1996). Structural basis for T cell recognition of altered peptide ligands: a single T cell receptor can productively recognize a large continuum of related ligands.(変化したペプチドリガンドのT細胞認識の構造的基礎:単一のT細胞レセプターは、関連するリガンドの大きな連続体を生産的に認識することができる)J Exp Med. 184, 1259-1268.
Le,B.N., Tan,A.T., Kunasegaran,K., Tham,C.Y.L., Hafezi,M., Chia,A., Chng,M.H.Y., Lin,M.,Tan,N., Linster,M., Chia,W.N., Chen,M.I., Wang,L.F., Ooi,E.E., Kalimuddin,S., Tambyah,P.A., Low,J.G., Tan,Y.J., and Bertoletti,A. (2020). SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls.(COVID-19およびSARS症例と非感染対照とにおける、SARS-CoV-2特異的T細胞免疫) Nature 584, 457-462.
Liu,W.J., Zhao,M., Liu,K., Xu,K., Wong,G., Tan,W., and Gao,G.F. (2017). T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV.(SARS-CoVのT細胞免疫:MERS-CoVに対するワクチン開発への示唆)Antiviral Res. 137, 82-92.
Mason,D. (1998). A very high level of crossreactivity is an essential feature of the T-cell receptor.(非常に高いレベルの交差反応性は、T細胞レセプターの本質的な特徴である。) Immunol. Today 19, 395-404.
Mateus,J., Grifoni,A., Tarke,A., Sidney,J., Ramirez,S.I., Dan,J.M., Burger,Z.C., Rawlings,S.A., Smith,D.M., Phillips,E., Mallal,S., Lammers,M., Rubiro,P., Quiambao,L., Sutherland,A., Yu,E.D., da Silva,A.R., Greenbaum,J., Frazier,A., Markmann,A.J., Premkumar,L., de,S.A., Peters,B., Crotty,S., Sette,A., and Weiskopf,D. (2020). Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans.(未感染のヒトにおけるSARS-CoV-2のT細胞エピトープの選択性と交差反応性) Science.
Petrosillo,N., Viceconte,G., Ergonul,O., Ippolito,G., and Petersen,E. (2020). COVID-19, SARS and MERS: are they closely related? (COVID-19、SARS、MERS:これらは密接に関連しているのか?)Clin Microbiol. Infect. 26, 729-734.
Rosenke,K., Meade-White,K., Letko,M., Clancy,C., Hansen,F., Liu,Y., Okumura,A., Tang-Huau,TL., Li,R., Saturday,G., Feldmann,F., Scott,D., Wang,Z., Munster,V., Jarvis,M.A., Feldmann,H. (2020). Defining the Syrian hamster as a highly susceptible preclinical model for SARS-CoV-2 infection.(シリアンハムスターをSARS-CoV-2感染の高感受性前臨床モデルとして定義) Emerg Microbes Infect. 9:2673-2684.
Severance,E.G., Bossis,I., Dickerson,F.B., Stallings,C.R., Origoni,A.E., Sullens,A., Yolken,R.H., and Viscidi,R.P. (2008). Development of a nucleocapsid-based human coronavirus immunoassay and estimates of individuals exposed to coronavirus in a U.S. metropolitan population.(ヌクレオカプシドベースのヒトコロナウイルス免疫測定法の開発と、米国大都市集団におけるコロナウイルス感染者の推定) Clin Vaccine Immunol. 15, 1805-1810.
Sewell,A.K. (2012). Why must T cells be cross-reactive?(なぜT細胞は交差反応性でなければならないのか?) Nat. Rev. Immunol. 12, 669-677.
Sun,S.H., Chen,Q., Gu,H.J., Yang,G., Wang,Y.X., Huang,X.Y., Liu,S.S., Zhang,N.N., Li,X.F., Xiong,R., Guo,Y., Deng,Y.Q., Huang,W.J., Liu,Q., Liu,Q.M., Shen,Y.L., Zhou,Y., Yang,X., Zhao,T.Y., Fan,C.F., Zhou,Y.S., Qin,C.F., and Wang,Y.C. (2020). A Mouse Model of SARS-CoV-2 Infection and Pathogenesis.(SARS-CoV-2感染および病態形成のマウスモデル) Cell Host. Microbe 28, 124-133.
Weiskopf,D., Schmitz,K.S., Raadsen,M.P., Grifoni,A., Okba,N.M.A., Endeman,H., van den Akker,J.P.C., Molenkamp,R., Koopmans,M.P.G., van Gorp,E.C.M., Haagmans,B.L., de Swart,R.L., Sette,A., and de Vries,R.D. (2020). Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome.(急性呼吸窮迫症候群のCOVID-19罹患者におけるSARS-CoV-2特異的T細胞の表現型と動態) Sci. Immunol. 5.
Wilson,D.B., Wilson,D.H., Schroder,K., Pinilla,C., Blondelle,S., Houghten,R.A., and Garcia,K.C. (2004). Specificity and degeneracy of T cells.(T細胞の特異性と変性) Mol. Immunol. 40, 1047-1055.
電子ファイルを添付した。
Claims (45)
- (a)キメラパピローマウイルス様粒子(VLP)であって、L1タンパク質と、前記L1タンパク質のHIループに挿入されたアミノ酸挿入物と、を含むVLPと、
(b)コロナウイルス抗原と、
を含む組成物。 - 前記アミノ酸挿入物は、負電荷アミノ酸の連続配列と末端システイン残基とを含む、
請求項1に記載の組成物。 - 前記負電荷アミノ酸は、グルタミン酸、アスパラギン酸、およびそれらの組み合わせからなる群から選択される、
請求項2に記載の組成物。 - 前記アミノ酸挿入物は、4~10の前記負電荷アミノ酸と、C末端、N末端、または両方における前記末端システイン残基と、を含む、
請求項2に記載の組成物。 - 前記アミノ酸挿入物は、表1に示される挿入物から選択される、
請求項2に記載の組成物。 - 前記アミノ酸挿入物は、344位と357位との間の位置で前記L1タンパク質の前記HIループに挿入されている、
請求項1に記載の組成物。 - 前記アミノ酸挿入物は、344位と357位との間の1以上の天然残基を置換する、
請求項6に記載の組成物。 - 前記アミノ酸挿入物は、表1に示される天然アミノ酸配列を置換する、
請求項6に記載の組成物。 - 前記コロナウイルス抗原は、OC43抗原、HKU1抗原、229E抗原、NL63抗原、SARS-CoV-1抗原、MERS抗原、SARS-CoV-2抗原、およびこれらの融合体からなる群から選択される、
請求項1に記載の組成物。 - 前記コロナウイルス抗原は、OC43、HKU1、229E、NL63、SARS-CoV-1、MERS、および/またはSARS-CoV-2のコロナウイルスに由来する1または複数のタンパク質を含む、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43、HKU1、229E、NL63、SARS-CoV-1、MERS、および/またはSARS-CoV-2のコロナウイルスに由来する、膜タンパク質(M)、ヌクレオカプシドタンパク質(N)、ならびにスパイク(S)エンベロープタンパク質のS2領域からなる群から選択される、ウイルス構造タンパク質を含む、請求項9に記載の組成物。
- 前記コロナウイルス抗原は、SARS-CoV-1またはSARS-CoV-2のコロナウイルスの、ORF3aタンパク質およびORF7aタンパク質からなる群から選択されるウイルス構造タンパク質を含む、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43、HKU1、229E、NL63、SARS-CoV-1、MERS、および/またはSARS-CoV-2コロナウイルスに由来する、nsp3、nsp4、nsp6、nsp7、およびnsp12タンパク質からなる群から選択されるウイルス非構造タンパク質を含む、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、膜タンパク質(M)、ヌクレオカプシドタンパク質(N)、スパイク(S)エンベロープタンパク質のS2領域、ORF3a、およびORF7aからなる群から選択されるSARS-CoV-2のウイルス構造タンパク質を含む、
請求項9に記載の組成物。 - 前記コロナウイルス構造タンパク質抗原は、表3~7のアミノ酸配列の1以上から選択される、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、SARS-CoV-2のnsp6、nsp7および/またはnsp12タンパク質に由来するウイルス非構造タンパク質を含む、請求項9に記載の組成物。
- 前記コロナウイルス非構造タンパク質抗原は、表8~10のアミノ酸配列の1以上から選択される、請求項9に記載の組成物。
- 前記コロナウイルス抗原は、膜タンパク質(M)、ヌクレオカプシドタンパク質(N)、およびスパイク(S)エンベロープタンパク質のS2領域からなる群から選択される、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1または複数のウイルス構造タンパク質を含む、請求項9に記載の組成物。
- 前記コロナウイルス抗原は、nsp3、nsp4、nsp6、nsp7および/またはnsp12タンパク質からなる群から選択される、OC43コロナウイルスの1以上のウイルス非構造タンパク質を含む、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1以上のウイルスタンパク質を含み、
前記1以上のウイルスタンパク質は、SARS-CoV-1コロナウイルス、MERSコロナウイルス、および/またはSARS-CoV-2コロナウイルスのアミノ酸配列と40%以上の同一性を有する、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1以上のウイルスタンパク質を含み、
前記1以上のウイルスタンパク質は、SARS-CoV-1コロナウイルス、MERSコロナウイルス、および/またはSARS-CoV-2コロナウイルスのアミノ酸配列と50%以上の同一性を有する、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1以上のウイルスタンパク質を含み、
前記1以上のウイルスタンパク質は、SARS-CoV-1コロナウイルス、MERSコロナウイルス、および/またはSARS-CoV-2コロナウイルスのアミノ酸配列と60%以上の同一性を有する、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1以上のウイルスタンパク質を含み、
前記1以上のウイルスタンパク質は、SARS-CoV-1コロナウイルス、MERSコロナウイルス、および/またはSARS-CoV-2コロナウイルスのアミノ酸配列と70%以上の同一性を有する、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、OC43コロナウイルスおよび/またはHKU1コロナウイルスの1以上のウイルスタンパク質を含み、
前記1以上のウイルスタンパク質は、SARS-CoV-1コロナウイルス、MERSコロナウイルス、および/またはSARS-CoV-2コロナウイルスのアミノ酸配列と80%以上の同一性を有する、
請求項9に記載の組成物。 - 前記OC43および/またはHKU1コロナウイルス構造タンパク質抗原は、表11~13のアミノ酸配列の1または複数から選択される、
請求項9に記載の組成物。 - 前記OC43コロナウイルス非構造抗原は、表14~17のアミノ酸配列の1以上から選択される、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、ウイルスタンパク質の抗原領域の全アミノ酸配列を含むポリペプチドのセットを含む、
請求項9に記載の組成物。 - 前記コロナウイルス抗原は、
OC43および/またはHKU1コロナウイルスの構造タンパク質および非構造タンパク質およびペプチドから選択される第1の抗原と、
SARS-CoV-2コロナウイルスの構造タンパク質および非構造タンパク質およびペプチドから選択される第2の抗原と、
を含む、請求項9に記載の組成物。 - 前記コロナウイルス抗原は、
OC43および/またはHKU1コロナウイルスの構造タンパク質および構造ペプチドから選択される第1の抗原と、
OC43の非構造タンパク質および非構造ペプチドから選択される第2の抗原と、
SARS-CoV-2コロナウイルスの構造タンパク質および構造ペプチドから選択される第3の抗原と、
SARS-CoV-2の非構造タンパク質および非構造ペプチドから選択される第4の抗原と、
を含む、請求項9に記載の組成物。 - 第1の末端で前記アミノ酸挿入物に連結され、第2の末端で前記コロナウイルス抗原に連結されるTAG配列をさらに含む、
請求項1に記載の組成物。 - 前記TAG配列は、ジスルフィド結合によってキメラパピローマウイルスに連結され、ペプチド結合によって前記コロナウイルス抗原に連結される、
請求項30に記載の組成物。 - 前記TAG配列は、C末端タンパク質プロセシング配列AAYYを含む、
請求項30に記載の組成物。 - 前記TAG配列は、4~10の正電荷アミノ酸および末端システイン残基の配列を含む、請求項30に記載の組成物。
- 前記TAG配列は、表2に示されるアミノ酸配列を含む、
請求項30に記載の組成物。 - 哺乳動物における細胞傷害性T細胞応答を刺激するのに有効である、
請求項1に記載の組成物。 - 哺乳動物におけるコロナウイルスに対する細胞傷害性T細胞応答を刺激するための方法であって、
請求項1~35のいずれか1項に記載の組成物を前記哺乳動物に投与すること、
を含む、方法。 - 前記組成物は、OC43抗原、HKU1抗原、229E抗原、NL63抗原、およびこれらの組み合わせからなる群から選択される、コロナウイルス抗原を含む、
請求項36に記載の方法。 - 前記コロナウイルスは、SARS-CoV-1、MERS、およびSARS-CoV-2のうちの1以上である、
請求項36に記載の方法。 - 哺乳動物における2以上のコロナウイルスに対する細胞傷害性T細胞応答を刺激するための方法であって、
請求項1~6のいずれか1項に記載の組成物を前記哺乳動物に投与すること、
を含み、
前記コロナウイルス抗原は、前記2以上のコロナウイルスのそれぞれに由来する少なくとも1つの抗原を含む、
方法。 - 前記組成物は、
OC43、HKU1、229EおよびNL63の構造タンパク質および構造ペプチドからなる群から選択される第1のコロナウイルス抗原と、
OC43、HKU1、229EおよびNL63の非構造タンパク質および非構造ペプチドからなる群から選択される第2のコロナウイルス抗原と、
SARS-CoV-1、MERSおよびSARS-CoV-2の構造タンパク質および構造ペプチドからなる群から選択される第3のコロナウイルス抗原と、
SARS-CoV-1、MERSおよびSARS-CoV-2の非構造タンパク質および非構造ペプチドからなる群から選択される第4のコロナウイルス抗原と、
を含む、請求項39に記載の方法。 - 前記細胞傷害性T細胞応答は、交差反応性記憶T細胞の刺激を含み、
前記交差反応性記憶T細胞は、前記第1のコロナウイルス抗原および前記第2のコロナウイルス抗原の投与によって刺激され、SARS-CoV-1、MERSおよびSARS-CoV-2のうちの1つに感染した細胞に対して細胞傷害性を示す、
請求項40に記載の方法。 - コロナウイルスに対する治療免疫および保護免疫の両方を刺激するための方法であって、
請求項1~35のいずれか1項に記載の組成物を対象に投与すること、
を含む、方法。 - 前記組成物は、
OC43、HKU1、229E、および/またはNL63の構造タンパク質または構造ペプチドからなる群から選択される第1のコロナウイルス抗原と、
OC43、HKU1、229E、およびNL63の非構造タンパク質または非構造ペプチドからなる群から選択される第2のコロナウイルス抗原と、
SARS-CoV-1、MERSおよびSARS-CoV-2の構造タンパク質および構造ペプチドからなる群から選択される第3のコロナウイルス抗原と、
SARS-CoV-1、MERSおよびSARS-CoV-2の非構造タンパク質および非構造ペプチドからなる群から選択される第4のコロナウイルス抗原と、
を含む、請求項42に記載の方法。 - 前記細胞傷害性T細胞応答は、交差反応性記憶T細胞の刺激を含み、
前記交差反応性記憶T細胞は、前記第1のコロナウイルス抗原および第2のコロナウイルス抗原の投与によって刺激され、SARS-CoV-1、MERSおよびSARS-CoV-2のうちの1つに感染した細胞に対して細胞傷害性を示す、
請求項43に記載の方法。 - 哺乳動物において事前に存在する交差反応性記憶T細胞を刺激するための方法であって、
請求項1~6のいずれか1項に記載の組成物を前記哺乳動物に投与することを含み、
事前に存在する前記交差反応性記憶T細胞は、OC43、HKU1、229E、およびNL63のうちの1つに対する感染に応答して誘導されたものであり、
前記交差反応性記憶T細胞は、SARS-CoV-1、MERS、およびSARS-CoV-2のうちの1つに感染した細胞に対して細胞傷害性を示す、
方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US202163137402P | 2021-01-14 | 2021-01-14 | |
| US63/137,402 | 2021-01-14 | ||
| PCT/US2022/012434 WO2022155417A2 (en) | 2021-01-14 | 2022-01-14 | Cytolytic t cell immunotherapy for highly pathogenic coronaviruses |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2024502837A true JP2024502837A (ja) | 2024-01-23 |
Family
ID=82447540
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2023541012A Pending JP2024502837A (ja) | 2021-01-14 | 2022-01-14 | 高病原性コロナウイルスに対する細胞傷害性t細胞免疫療法 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US20250276056A1 (ja) |
| EP (1) | EP4277657A4 (ja) |
| JP (1) | JP2024502837A (ja) |
| KR (1) | KR20230131871A (ja) |
| CN (1) | CN117157310A (ja) |
| AU (1) | AU2022208375A1 (ja) |
| CA (1) | CA3204663A1 (ja) |
| IL (1) | IL304175A (ja) |
| MX (1) | MX2023008242A (ja) |
| WO (1) | WO2022155417A2 (ja) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN113980099B (zh) * | 2021-03-29 | 2025-06-13 | 军事科学院军事医学研究院生命组学研究所 | 新冠病毒的n抗原特定表位及其应用 |
| GB202313360D0 (en) * | 2023-09-01 | 2023-10-18 | NEC Oncolmmunity AS | T cell broad betacoronavirus vaccine |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20170327543A1 (en) * | 2010-09-08 | 2017-11-16 | The Johns Hopkins University | Polyionic papilloma virus-like particle (vlp) vaccines |
| US20200061185A1 (en) * | 2016-10-25 | 2020-02-27 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Servic | Prefusion coronavirus spike proteins and their use |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| BR112017003462B1 (pt) * | 2014-09-03 | 2023-02-14 | Intervet International B.V. | Coronavírus bovino atenuado e vacina |
| EP4011451A1 (en) * | 2015-10-22 | 2022-06-15 | ModernaTX, Inc. | Metapneumovirus mrna vaccines |
| AU2019414934A1 (en) * | 2018-12-27 | 2021-06-24 | Verimmune Llc | Conjugated virus-like particles and uses thereof as anti-tumor immune redirectors |
| US11130787B2 (en) * | 2020-06-11 | 2021-09-28 | MBF Therapeutics, Inc. | Alphaherpesvirus glycoprotein d-encoding nucleic acid constructs and methods |
-
2022
- 2022-01-14 JP JP2023541012A patent/JP2024502837A/ja active Pending
- 2022-01-14 AU AU2022208375A patent/AU2022208375A1/en not_active Abandoned
- 2022-01-14 CN CN202280009729.8A patent/CN117157310A/zh active Pending
- 2022-01-14 MX MX2023008242A patent/MX2023008242A/es unknown
- 2022-01-14 US US18/272,331 patent/US20250276056A1/en active Pending
- 2022-01-14 KR KR1020237025487A patent/KR20230131871A/ko active Pending
- 2022-01-14 CA CA3204663A patent/CA3204663A1/en active Pending
- 2022-01-14 WO PCT/US2022/012434 patent/WO2022155417A2/en not_active Ceased
- 2022-01-14 EP EP22740111.4A patent/EP4277657A4/en not_active Withdrawn
-
2023
- 2023-07-02 IL IL304175A patent/IL304175A/en unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20170327543A1 (en) * | 2010-09-08 | 2017-11-16 | The Johns Hopkins University | Polyionic papilloma virus-like particle (vlp) vaccines |
| US20200061185A1 (en) * | 2016-10-25 | 2020-02-27 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Servic | Prefusion coronavirus spike proteins and their use |
Non-Patent Citations (3)
| Title |
|---|
| GRIFONI ALBA ET AL.: "Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed", CELL, vol. 181, JPN6025041606, 25 June 2020 (2020-06-25), pages 1489 - 1501, ISSN: 0005834434 * |
| ONG EDISON ET AL.: "COVID-19 Coronavirus Vaccine Design Using Reverse Vaccinology and Machine Learning", FRONTIERS IN IMMUNOLOGY, vol. Vol.11, Article 1581, JPN6025041604, 3 July 2020 (2020-07-03), pages 1 - 13, ISSN: 0005834432 * |
| SAFAVI ASHKAN ET AL.: "Exploring the out of sight antigens of SARS-CoV-2 to design a candidate multi-epitope vaccine by uti", VACCINE, vol. 38, JPN6025041605, 9 October 2020 (2020-10-09), pages 7612 - 7628, ISSN: 0005834433 * |
Also Published As
| Publication number | Publication date |
|---|---|
| US20250276056A1 (en) | 2025-09-04 |
| CN117157310A (zh) | 2023-12-01 |
| KR20230131871A (ko) | 2023-09-14 |
| EP4277657A2 (en) | 2023-11-22 |
| AU2022208375A9 (en) | 2024-10-24 |
| IL304175A (en) | 2023-09-01 |
| CA3204663A1 (en) | 2022-07-21 |
| MX2023008242A (es) | 2023-07-26 |
| WO2022155417A2 (en) | 2022-07-21 |
| EP4277657A4 (en) | 2025-06-04 |
| WO2022155417A3 (en) | 2022-08-25 |
| AU2022208375A1 (en) | 2023-07-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN111088283B (zh) | mVSV病毒载体及其病毒载体疫苗、一种基于mVSV介导的新冠肺炎疫苗 | |
| CN111218459B (zh) | 一种以人复制缺陷腺病毒为载体的重组新型冠状病毒疫苗 | |
| CN113666990B (zh) | 一种诱导广谱抗冠状病毒的t细胞疫苗免疫原及其应用 | |
| CN114096675A (zh) | 冠状病毒免疫原性组合物和其用途 | |
| JP2023513913A (ja) | 麻疹ベクターを用いたcovid-19免疫原性組成物及びワクチン | |
| EP3394085B1 (en) | Feline calicivirus vaccine | |
| JP2003180385A (ja) | コロナウイルスに対するワクチン接種用組成物および方法 | |
| CN102497880A (zh) | 新的人乳头状瘤病毒(hpv)蛋白构建体及其在预防hpv疾病中的用途 | |
| CN105980570A (zh) | Cmv疫苗 | |
| JPH08501931A (ja) | コロナウイルスに対するワクチン接種用組成物および方法 | |
| WO2023023940A1 (zh) | 一种诱导广谱抗冠状病毒的t细胞疫苗免疫原及其应用 | |
| JP2024502837A (ja) | 高病原性コロナウイルスに対する細胞傷害性t細胞免疫療法 | |
| US20210401983A1 (en) | Arthrogenic alphavirus vaccine | |
| Li et al. | The CDE region of feline Calicivirus VP1 protein is a potential candidate subunit vaccine | |
| CN105085672B (zh) | 3d蛋白特异性单克隆免疫球蛋白a抗体及其组合物 | |
| US20080069830A1 (en) | Dna Sequences, Peptides, Antibodies and Vaccines for Prevention and Treatment of Sars | |
| US20250312441A1 (en) | Immunogenic compositions for herpes simplex virus proteins | |
| ES2339728B1 (es) | Proteinas n, m y he de torovirus porcino, procedimiento de obtencion y sus aplicaciones en diagnostico y tratamiento de torovirus porcino. | |
| Navaratnarajah | Development of measles virus-based shielded oncolytic vectors: suitability of other paramyxovirus glycoproteins | |
| CN120442567A (zh) | 狂犬病毒糖蛋白嵌合重组水泡性口炎病毒及其应用 | |
| US20100317567A1 (en) | Diagnosing and protecting horses against papillomavirus | |
| HK1230514B (zh) | Pres1用於制备乙肝疫苗以及治疗慢性乙型肝炎的用途 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20241115 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20251007 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20260407 |